

Rapid production of new oligodendrocytes is required in the earliest stages of motor-skill learning
- PMID: 27455109
- PMCID: PMC5008443
- DOI: 10.1038/nn.4351
Rapid production of new oligodendrocytes is required in the earliest stages of motor-skill learning
Abstract
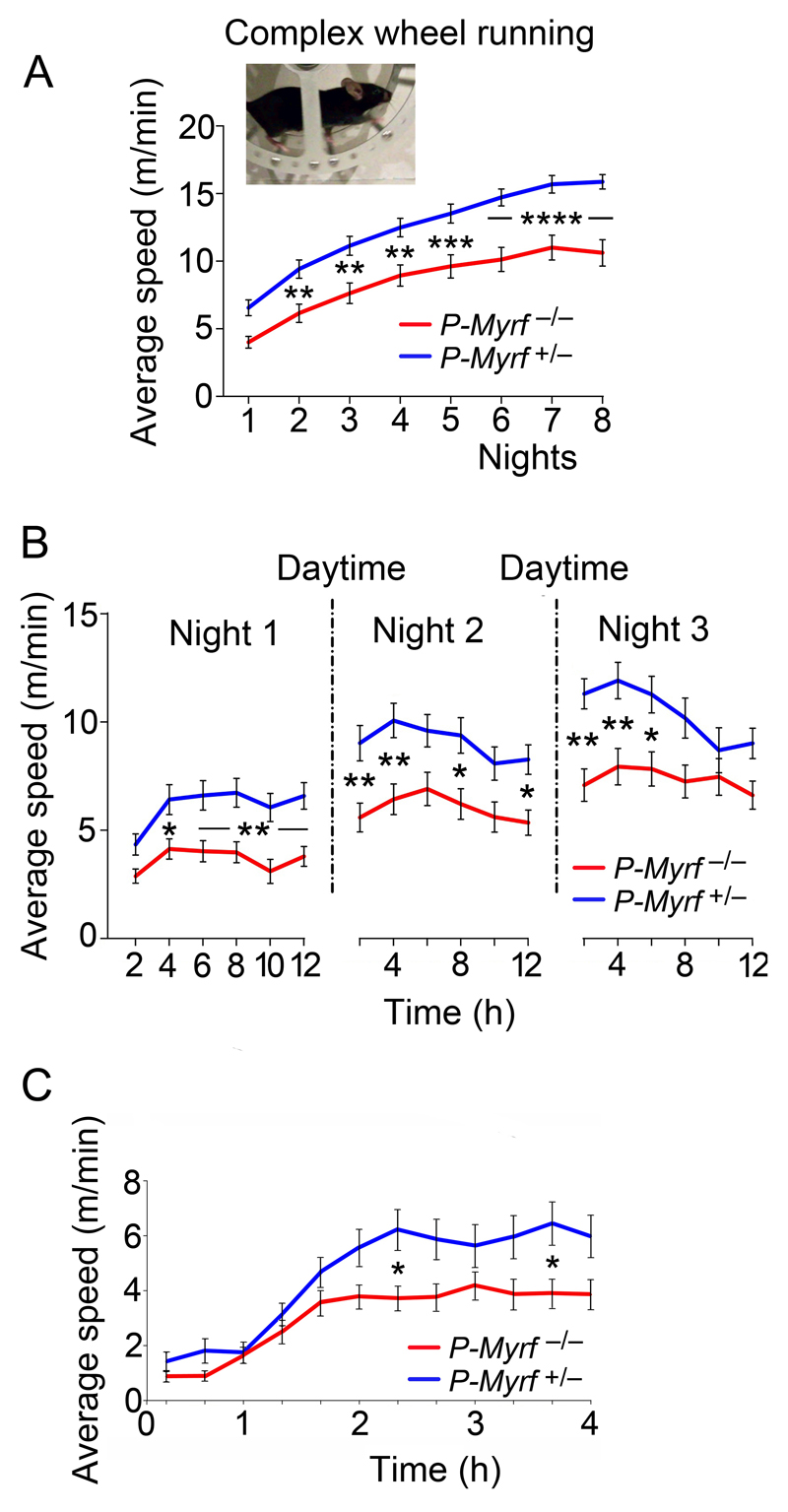
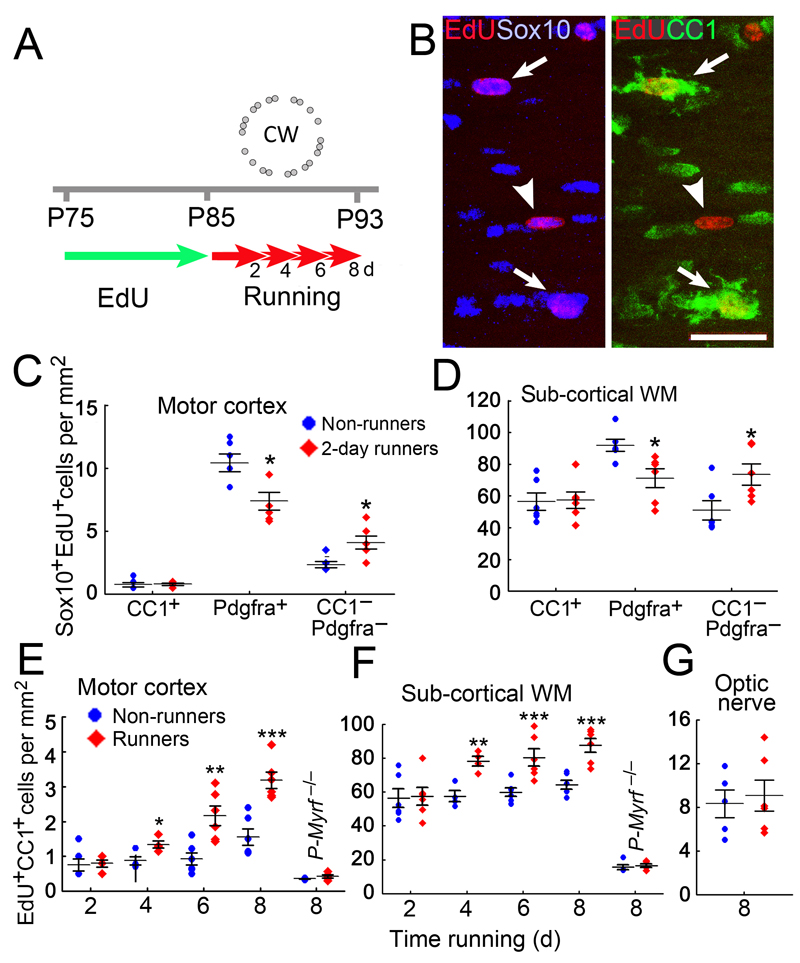
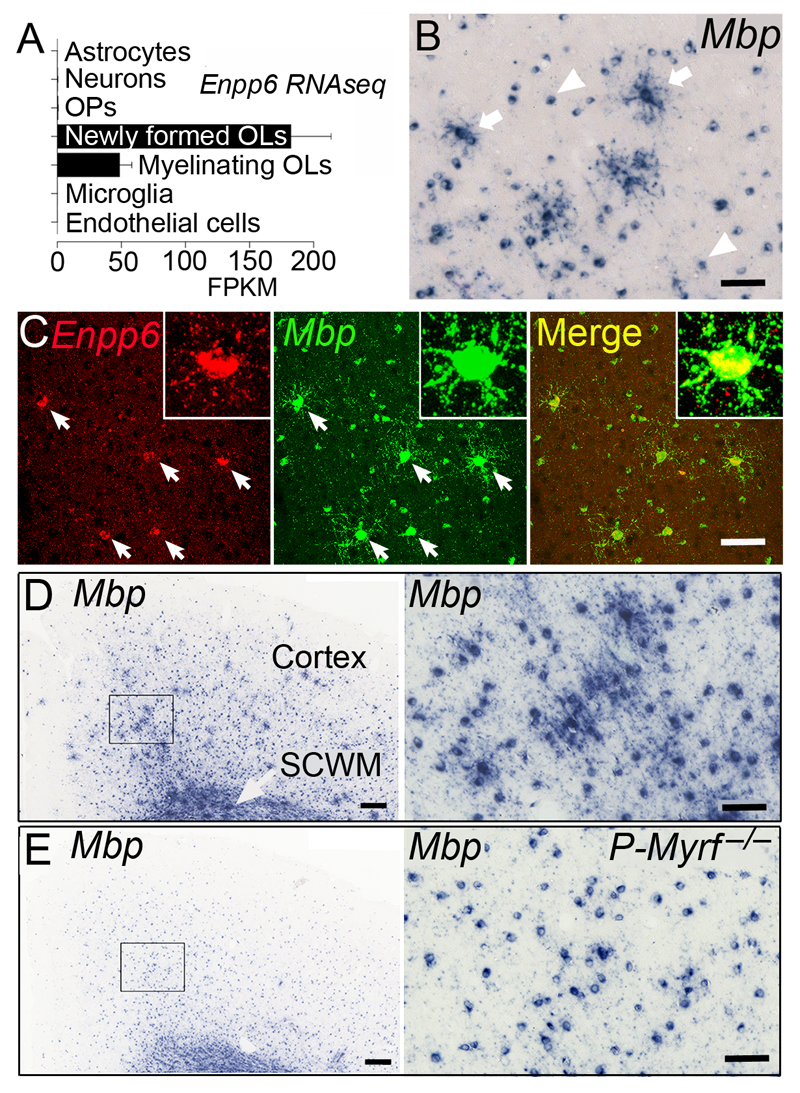
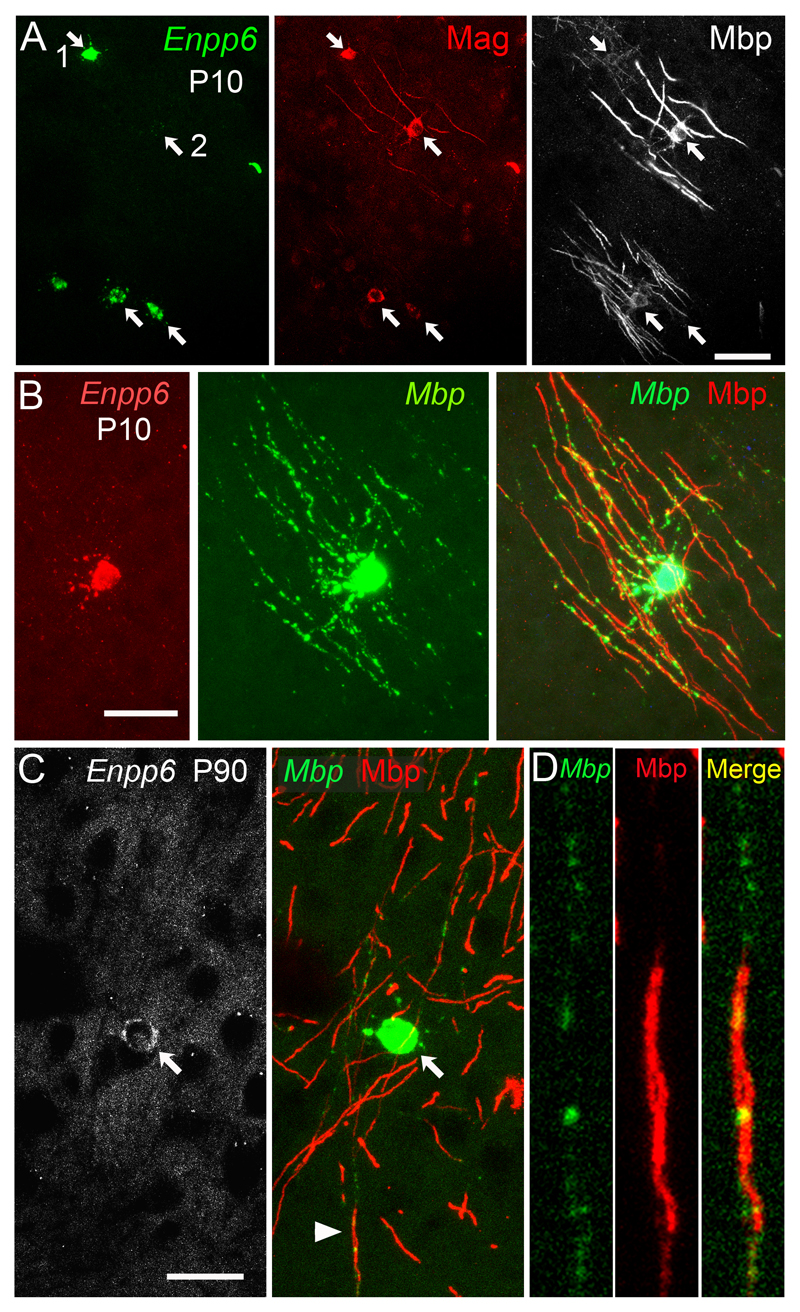
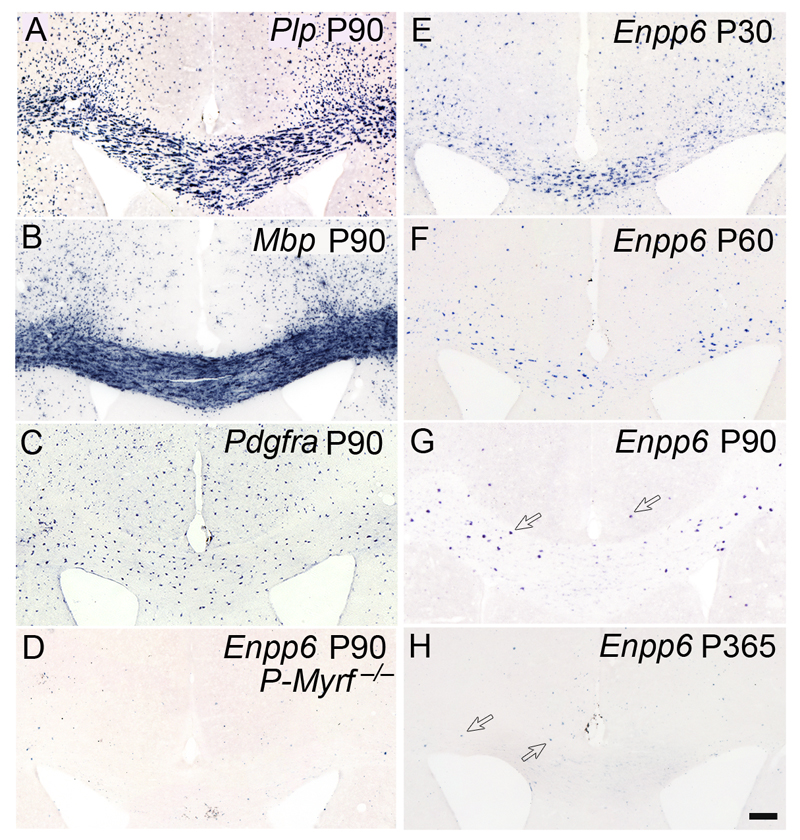
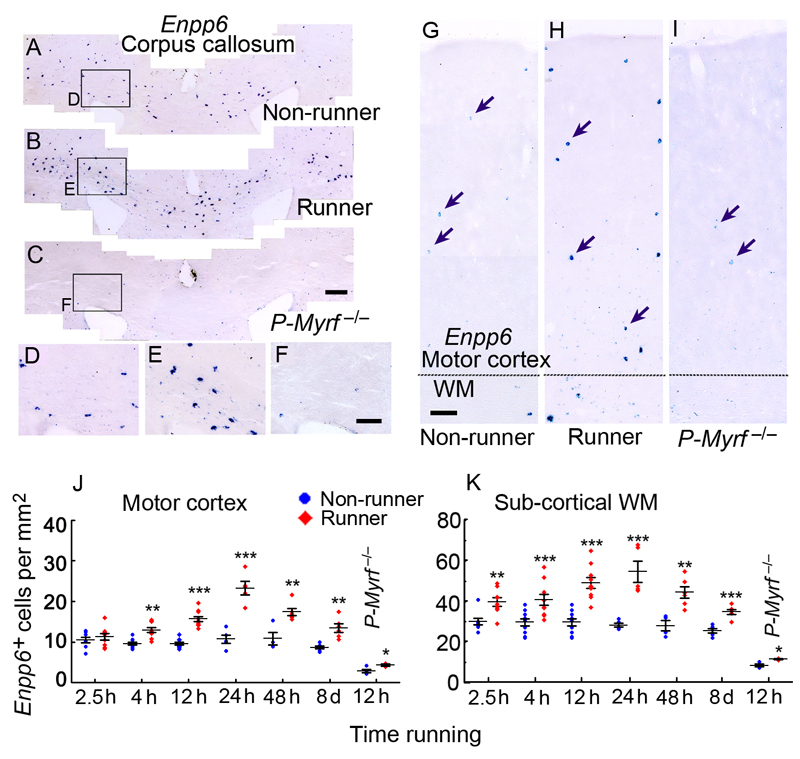
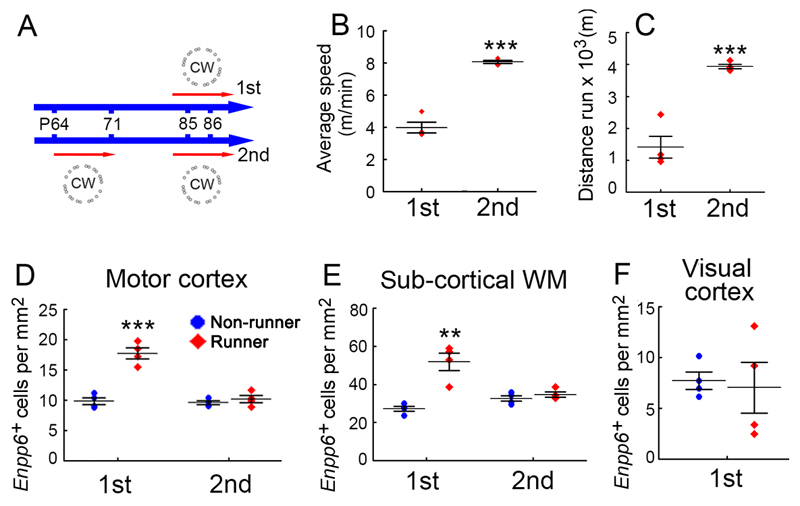
We identified mRNA encoding the ecto-enzyme Enpp6 as a marker of newly forming oligodendrocytes, and used Enpp6 in situ hybridization to track oligodendrocyte differentiation in adult mice as they learned a motor skill (running on a wheel with unevenly spaced rungs). Within just 2.5 h of exposure to the complex wheel, production of Enpp6-expressing immature oligodendrocytes was accelerated in subcortical white matter; within 4 h, it was accelerated in motor cortex. Conditional deletion of myelin regulatory factor (Myrf) in oligodendrocyte precursors blocked formation of new Enpp6(+) oligodendrocytes and impaired learning within the same ∼2-3 h time frame. This very early requirement for oligodendrocytes suggests a direct and active role in learning, closely linked to synaptic strengthening. Running performance of normal mice continued to improve over the following week accompanied by secondary waves of oligodendrocyte precursor proliferation and differentiation. We concluded that new oligodendrocytes contribute to both early and late stages of motor skill learning.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
