

Viperin inhibits rabies virus replication via reduced cholesterol and sphingomyelin and is regulated upstream by TLR4
- PMID: 27456665
- PMCID: PMC4960569
- DOI: 10.1038/srep30529
Viperin inhibits rabies virus replication via reduced cholesterol and sphingomyelin and is regulated upstream by TLR4
Abstract
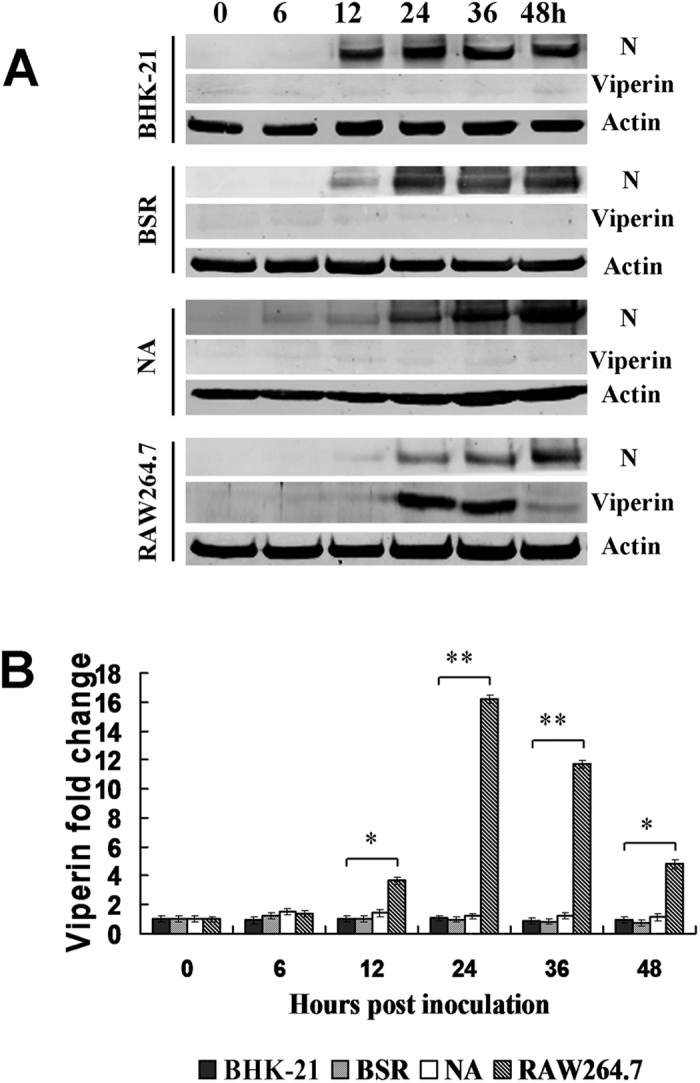
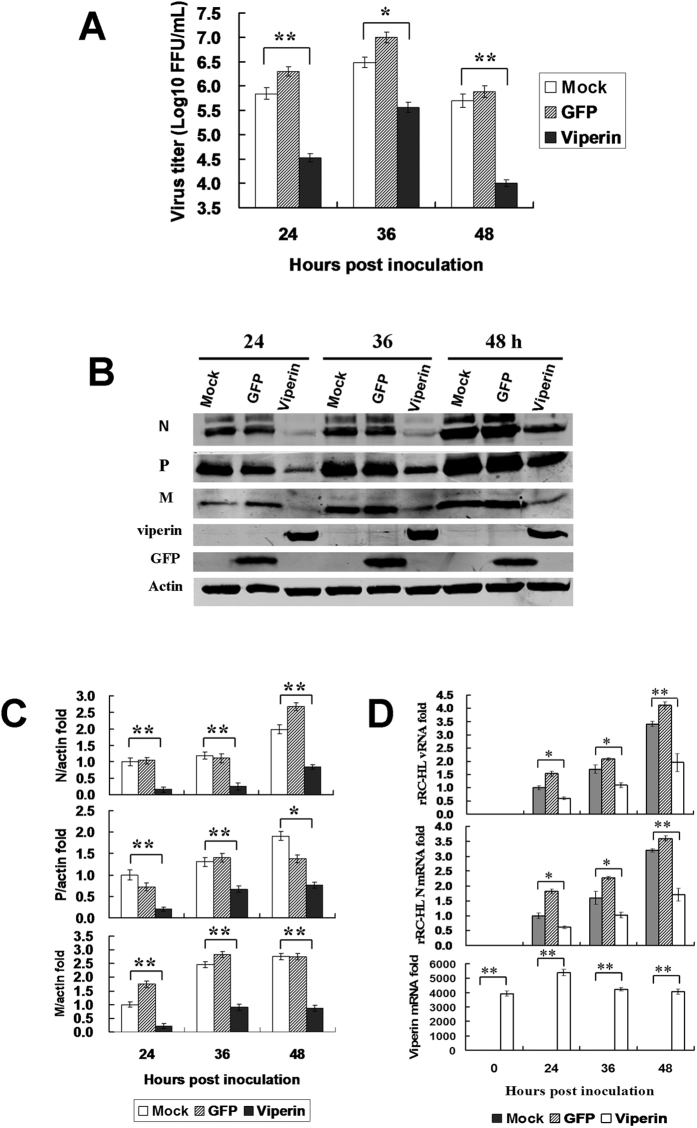
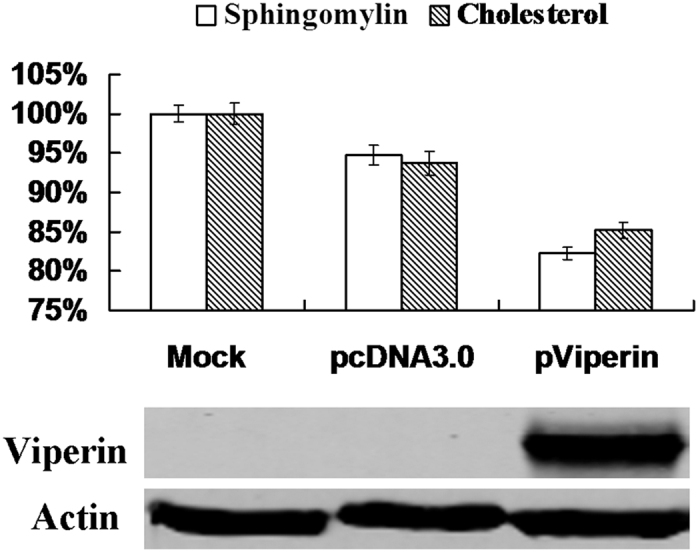
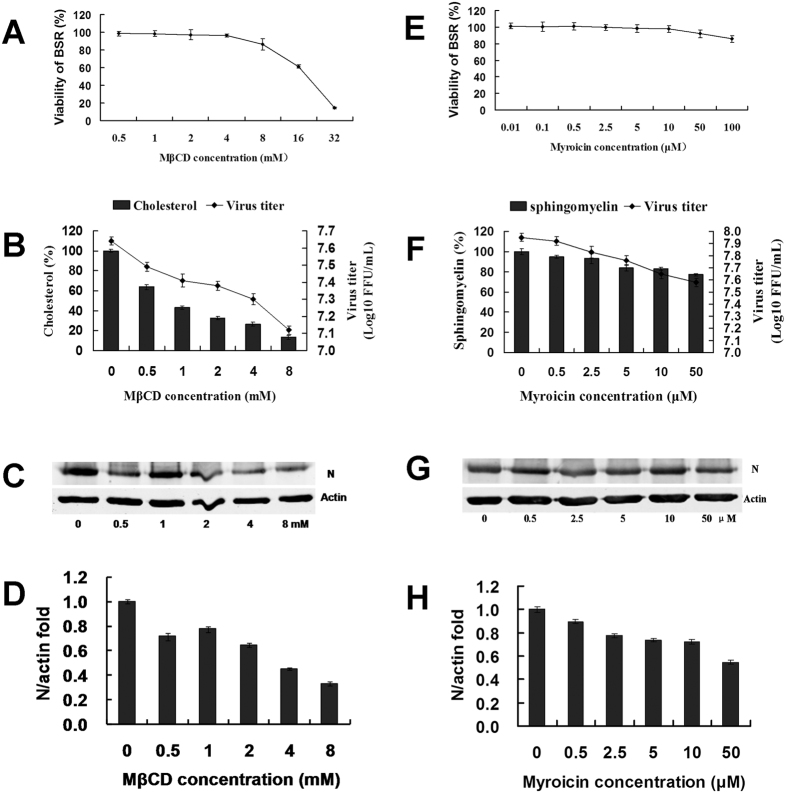
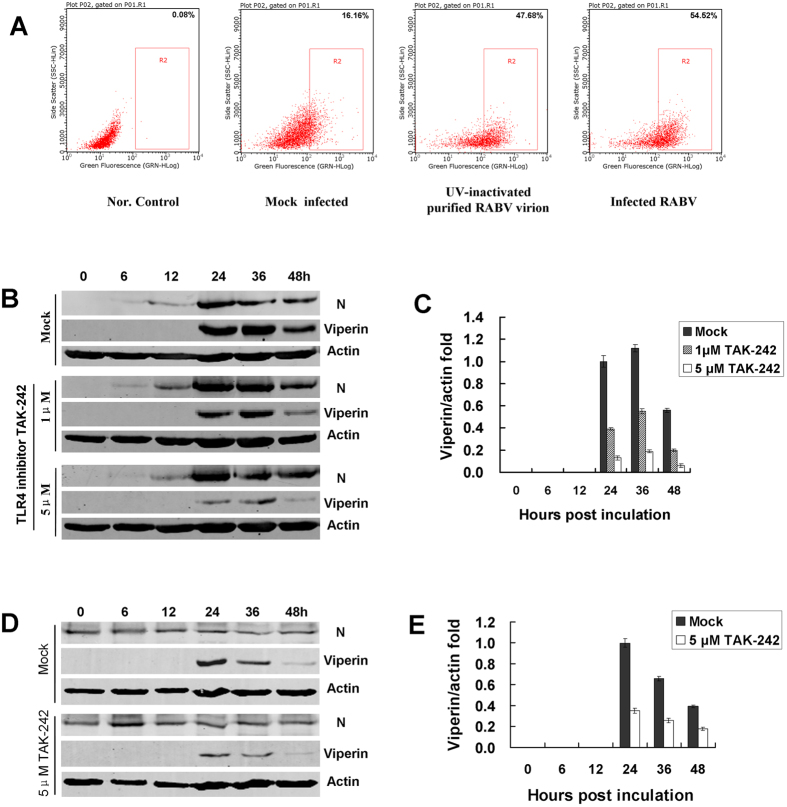
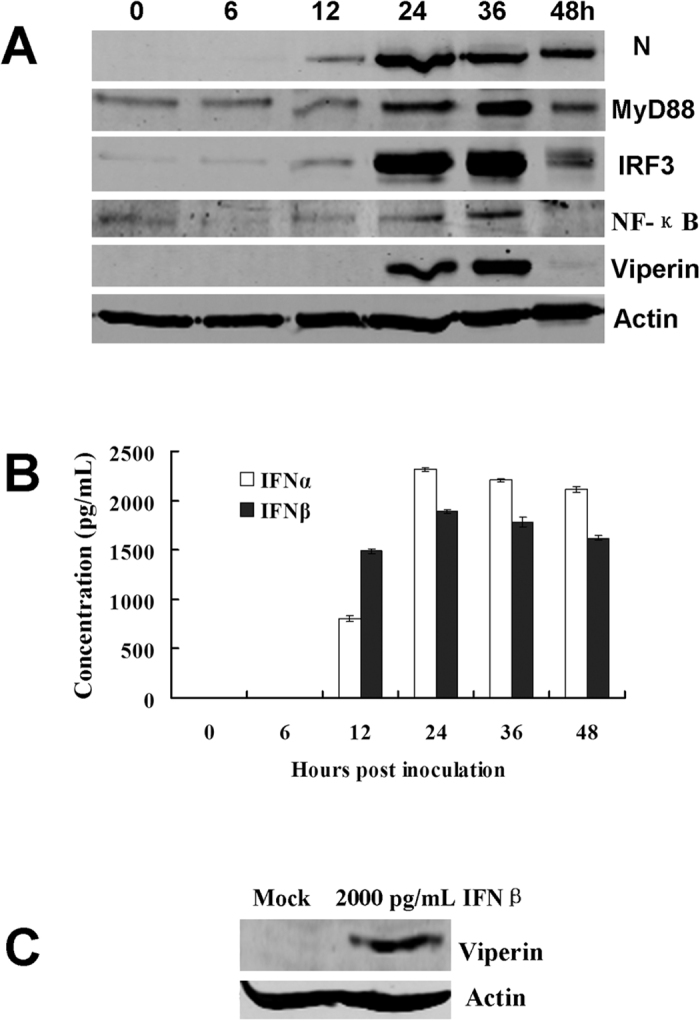
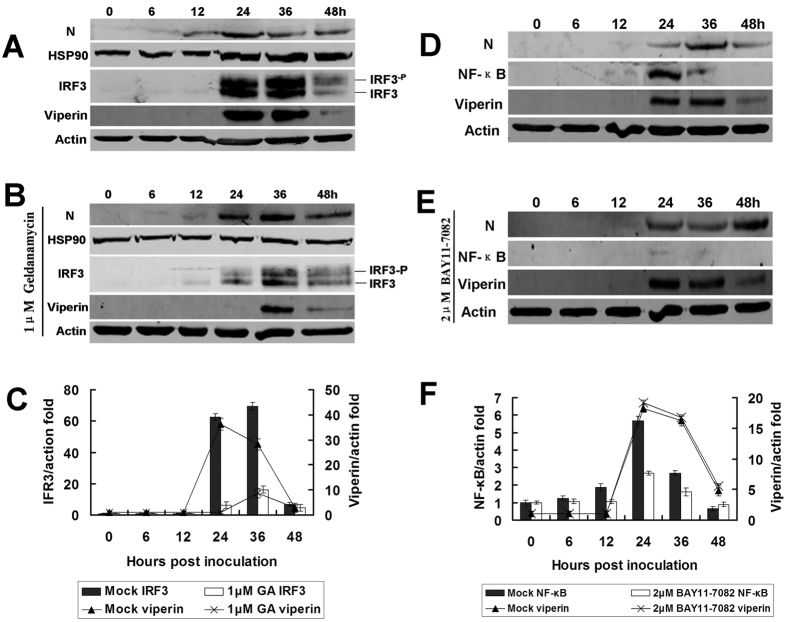
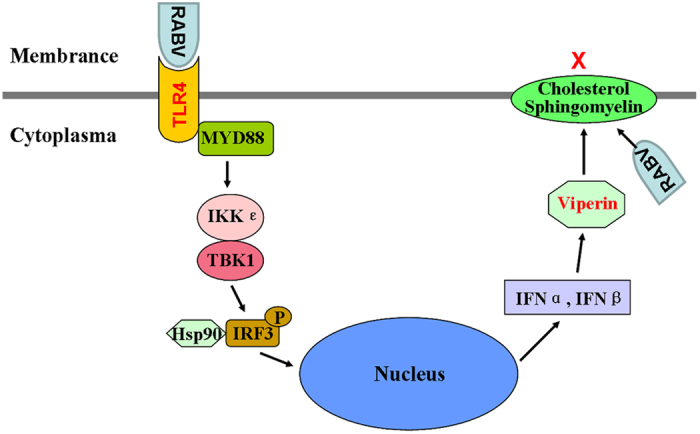
Viperin (virus inhibitory protein, endoplasmic reticulum-associated, IFN-inducible) is an interferon-inducible protein that mediates antiviral activity. Generally, rabies virus (RABV) multiplies extremely well in susceptible cells, leading to high virus titres. In this study, we found that viperin was significantly up-regulated in macrophage RAW264.7 cells but not in NA, BHK-21 or BSR cells. Transient viperin overexpression in BSR cells and stable expression in BHK-21 cells could inhibit RABV replication, including both attenuated and street RABV. Furthermore, the inhibitory function of viperin was related to reduce cholesterol/sphingomyelin on the membranes of RAW264.7 cells. We explored the up-stream regulation pathway of viperin in macrophage RAW264.7 cells in the context of RABV infection. An experiment confirmed that a specific Toll-like receptor 4 (TLR4) inhibitor, TAK-242, could inhibit viperin expression in RABV-infected RAW264.7 cells. These results support a regulatory role for TLR4. Geldanamycin, a specific inhibitor of interferon regulatory factor 3 (IRF3) (by inhibiting heat-shock protein 90 (Hsp90) of the IRF3 phosphorylation chaperone), significantly delayed and reduced viperin expression, indicating that IRF3 is involved in viperin induction in RAW264.7 cells. Taken together, our data support the therapeutic potential for viperin to inhibit RABV replication, which appears to involve upstream regulation by TLR4.
Figures
References
-
- Kawai T. & Akira S. Pathogen recognition with Toll-like receptors. Curr Opin Immunol. 17(4), 338–344 (2005). - PubMed
-
- Johnson N. et al. Lyssavirus infection activates interferon gene expression in the brain. J Gen Virol. 87, 2663–2667 (2006). - PubMed
-
- Saitoh T. et al. Antiviral protein Viperin promotes Toll-like receptor 7-and Toll-like receptor 9-mediated type I interferon production in plasmacytoid dendritic cells. Immunity. 34(3), 352–363 (2011). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
