

Avoidance and Subversion of Eukaryotic Homeostatic Autophagy Mechanisms by Bacterial Pathogens
- PMID: 27456933
- PMCID: PMC5010449
- DOI: 10.1016/j.jmb.2016.07.007
Avoidance and Subversion of Eukaryotic Homeostatic Autophagy Mechanisms by Bacterial Pathogens
Abstract
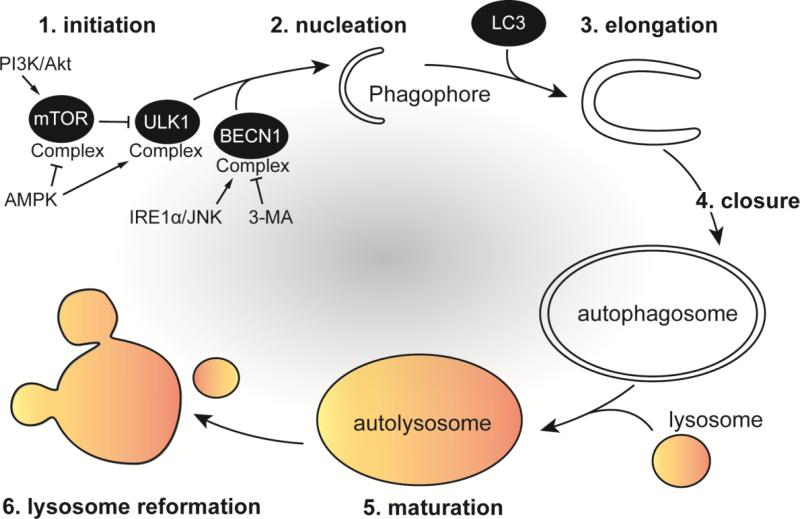
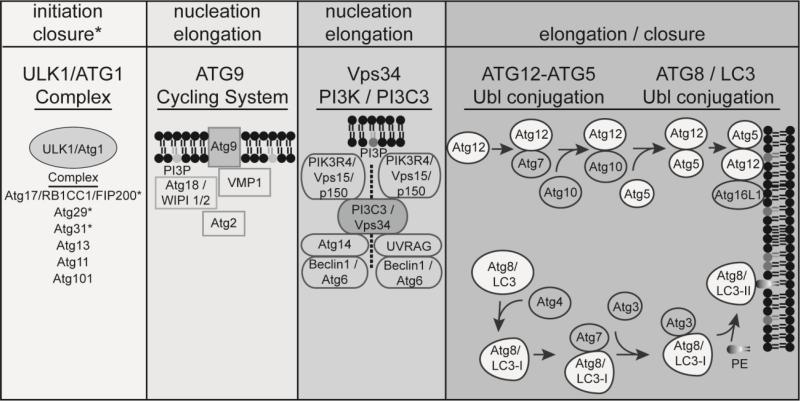
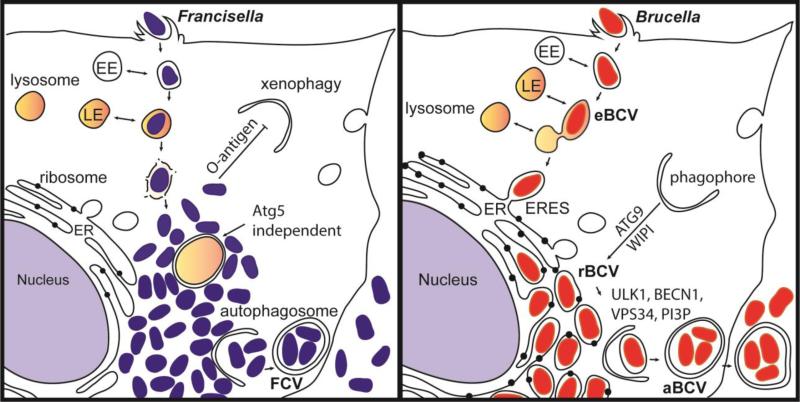
Autophagy is a conserved lysosomal recycling process, which maintains cellular homeostasis during stress and starvation conditions by degrading and recycling proteins, lipids, and carbohydrates, ultimately increasing nutrient availability in eukaryotes. An additional function of autophagy, termed xenophagy, is to detect, capture, and destroy invading microorganisms, such as viruses, bacteria, and protozoa, providing autophagy with a role in innate immunity. Many intracellular pathogens have, however, developed mechanisms to avoid xenophagy and have evolved strategies to take advantage of select autophagic processes to undergo their intracellular life cycle. This review article will discuss the molecular mechanisms used by the intracellular bacterial pathogens Francisella spp. and Brucella spp. to manipulate components of the autophagic pathway, promoting cytosolic growth in the case of Francisella spp. and facilitating cellular egress and cell-to-cell spread in the case of Brucella spp. These examples highlight how successful, highly infectious bacterial pathogens avoid or subvert host autophagy mechanisms normally employed to maintain eukaryotic homeostasis.
Keywords: O-antigen; bacterial secretion system; cellular egress; infection; nutrient acquisition.
Copyright © 2016 Elsevier Ltd. All rights reserved.
Figures
References
-
- Mizushima N. Autophagy: process and function. Genes Dev. 2007;21:2861–2873. doi:10.1101/gad.1599207. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
