

An Arginine Finger Regulates the Sequential Action of Asymmetrical Hexameric ATPase in the Double-Stranded DNA Translocation Motor
- PMID: 27457616
- PMCID: PMC5021374
- DOI: 10.1128/MCB.00142-16
An Arginine Finger Regulates the Sequential Action of Asymmetrical Hexameric ATPase in the Double-Stranded DNA Translocation Motor
Abstract
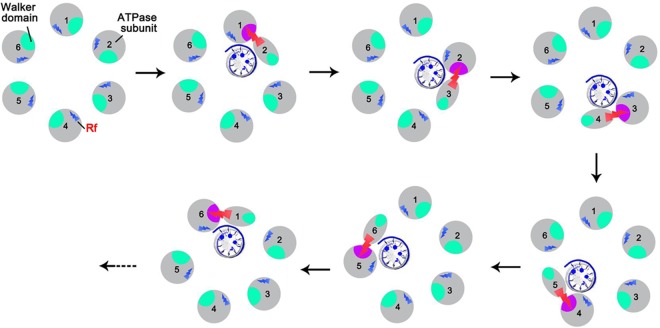
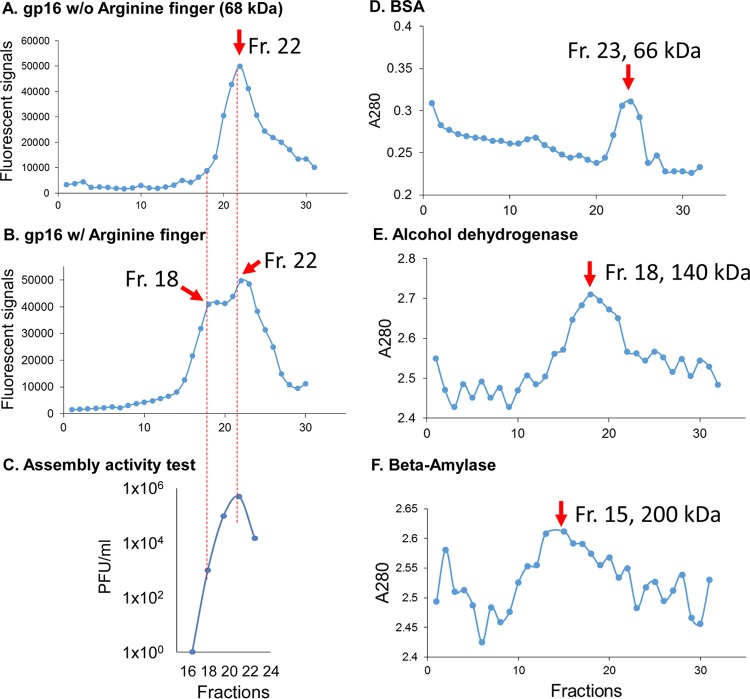
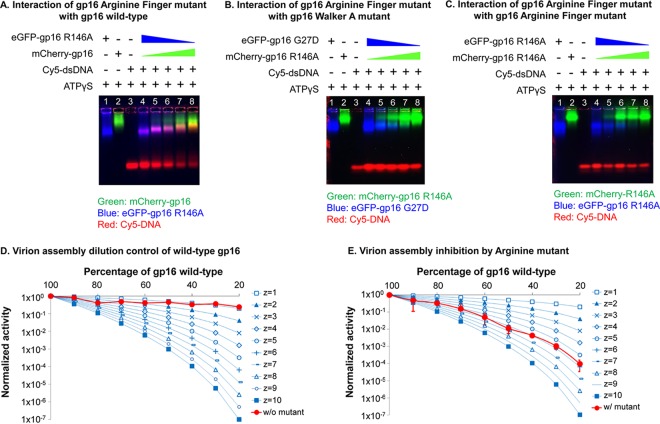
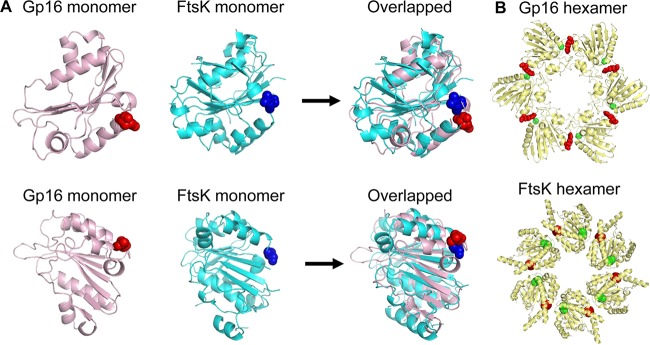
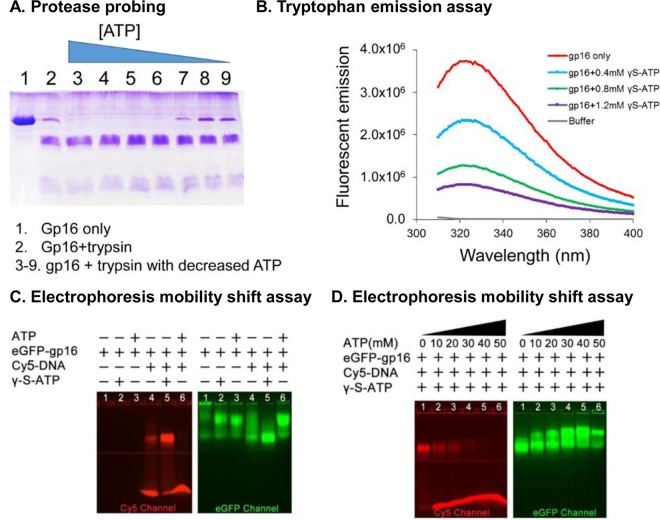
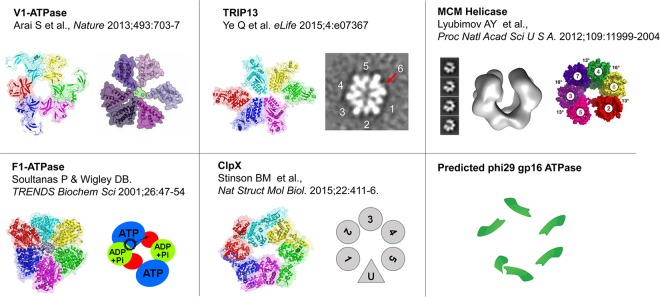
Biological motors are ubiquitous in living systems. Currently, how the motor components coordinate the unidirectional motion is elusive in most cases. Here, we report that the sequential action of the ATPase ring in the DNA packaging motor of bacteriophage ϕ29 is regulated by an arginine finger that extends from one ATPase subunit to the adjacent unit to promote noncovalent dimer formation. Mutation of the arginine finger resulted in the interruption of ATPase oligomerization, ATP binding/hydrolysis, and DNA translocation. Dimer formation reappeared when arginine mutants were mixed with other ATPase subunits that can offer the arginine to promote their interaction. Ultracentrifugation and virion assembly assays indicated that the ATPase was presenting as monomers and dimer mixtures. The isolated dimer alone was inactive in DNA translocation, but the addition of monomer could restore the activity, suggesting that the hexameric ATPase ring contained both dimer and monomers. Moreover, ATP binding or hydrolysis resulted in conformation and entropy changes of the ATPase with high or low DNA affinity. Taking these observations together, we concluded that the arginine finger regulates sequential action of the motor ATPase subunit by promoting the formation of the dimer inside the hexamer. The finding of asymmetrical hexameric organization is supported by structural evidence of many other ATPase systems showing the presence of one noncovalent dimer and four monomer subunits. All of these provide clues for why the asymmetrical hexameric ATPase gp16 of ϕ29 was previously reported as a pentameric configuration by cryo-electron microscopy (cryo-EM) since the contact by the arginine finger renders two adjacent ATPase subunits closer than other subunits. Thus, the asymmetrical hexamer would appear as a pentamer by cryo-EM, a technology that acquires the average of many images.
Copyright © 2016 Zhao et al.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources