

Reduced nighttime transpiration is a relevant breeding target for high water-use efficiency in grapevine
- PMID: 27457942
- PMCID: PMC4987834
- DOI: 10.1073/pnas.1600826113
Reduced nighttime transpiration is a relevant breeding target for high water-use efficiency in grapevine
Abstract
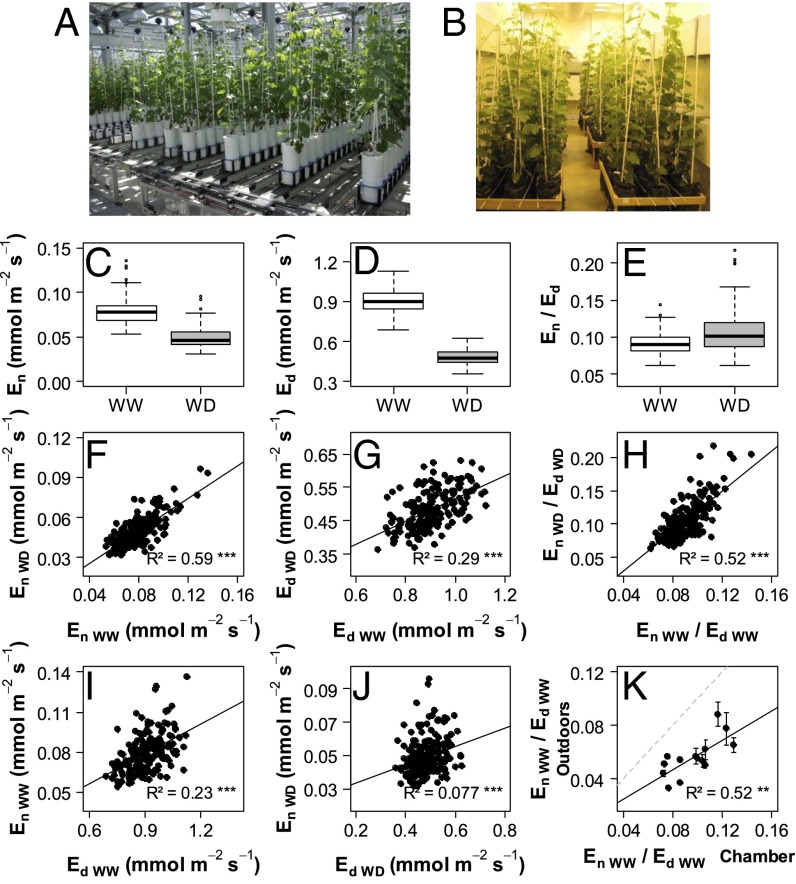
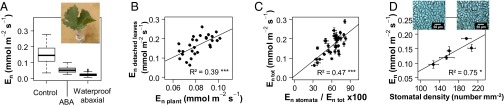
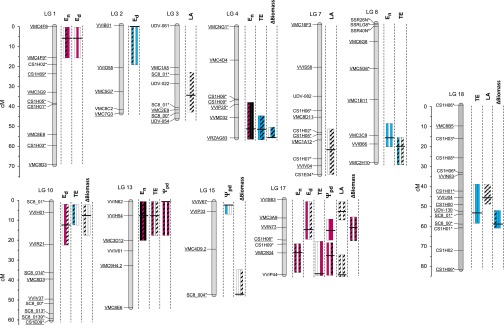
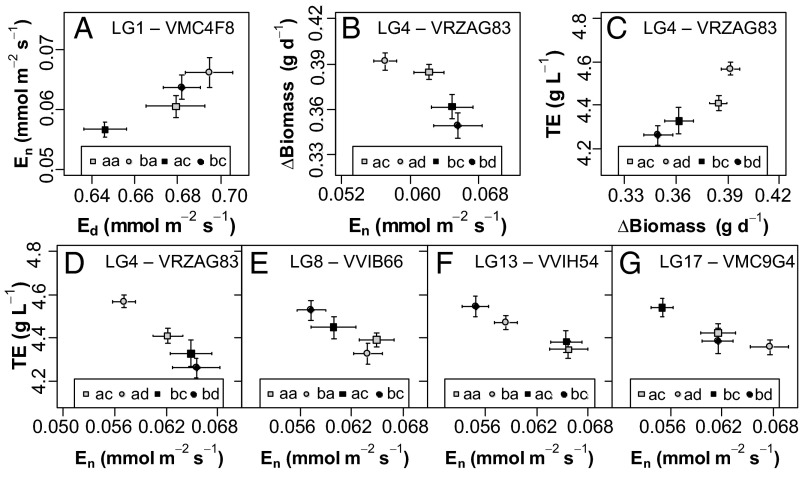
Increasing water scarcity challenges crop sustainability in many regions. As a consequence, the enhancement of transpiration efficiency (TE)-that is, the biomass produced per unit of water transpired-has become crucial in breeding programs. This could be achieved by reducing plant transpiration through a better closure of the stomatal pores at the leaf surface. However, this strategy generally also lowers growth, as stomatal opening is necessary for the capture of atmospheric CO2 that feeds daytime photosynthesis. Here, we considered the reduction in transpiration rate at night (En) as a possible strategy to limit water use without altering growth. For this purpose, we carried out a genetic analysis for En and TE in grapevine, a major crop in drought-prone areas. Using recently developed phenotyping facilities, potted plants of a cross between Syrah and Grenache cultivars were screened for 2 y under well-watered and moderate soil water deficit scenarios. High genetic variability was found for En under both scenarios and was primarily associated with residual diffusion through the stomata. Five quantitative trait loci (QTLs) were detected that underlay genetic variability in En Interestingly, four of them colocalized with QTLs for TE. Moreover, genotypes with favorable alleles on these common QTLs exhibited reduced En without altered growth. These results demonstrate the interest of breeding grapevine for lower water loss at night and pave the way to breeding other crops with this underexploited trait for higher TE.
Keywords: QTL; growth; night transpiration; stomata; transpiration efficiency.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Intergovernmental Panel on Climate Change . In: Climate Change 2013: The Physical Science Basis. Stocker TF, et al., editors. Cambridge Univ Press; New York: 2013.
-
- Condon AG, Richards RA, Rebetzke GJ, Farquhar GD. Breeding for high water-use efficiency. J Exp Bot. 2004;55(407):2447–2460. - PubMed
-
- Hetherington AM, Woodward FI. The role of stomata in sensing and driving environmental change. Nature. 2003;424(6951):901–908. - PubMed
-
- Bacon MA. What Is Water-Use Efficiency? Water Use Efficiency in Plant Biology. Blackwell; Oxford: 2004. pp. 27–41.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
