

Earth's oxygen cycle and the evolution of animal life
- PMID: 27457943
- PMCID: PMC4987840
- DOI: 10.1073/pnas.1521544113
Earth's oxygen cycle and the evolution of animal life
Abstract
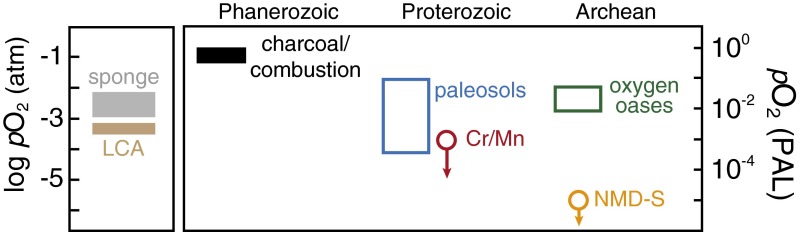
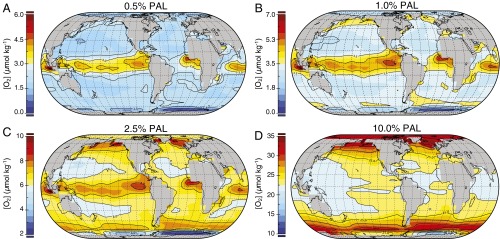
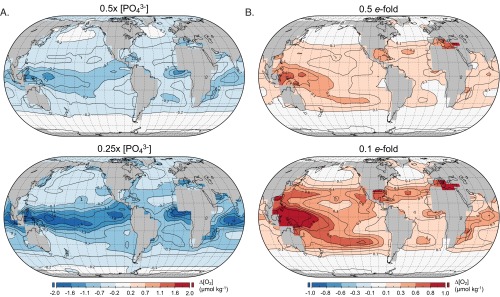
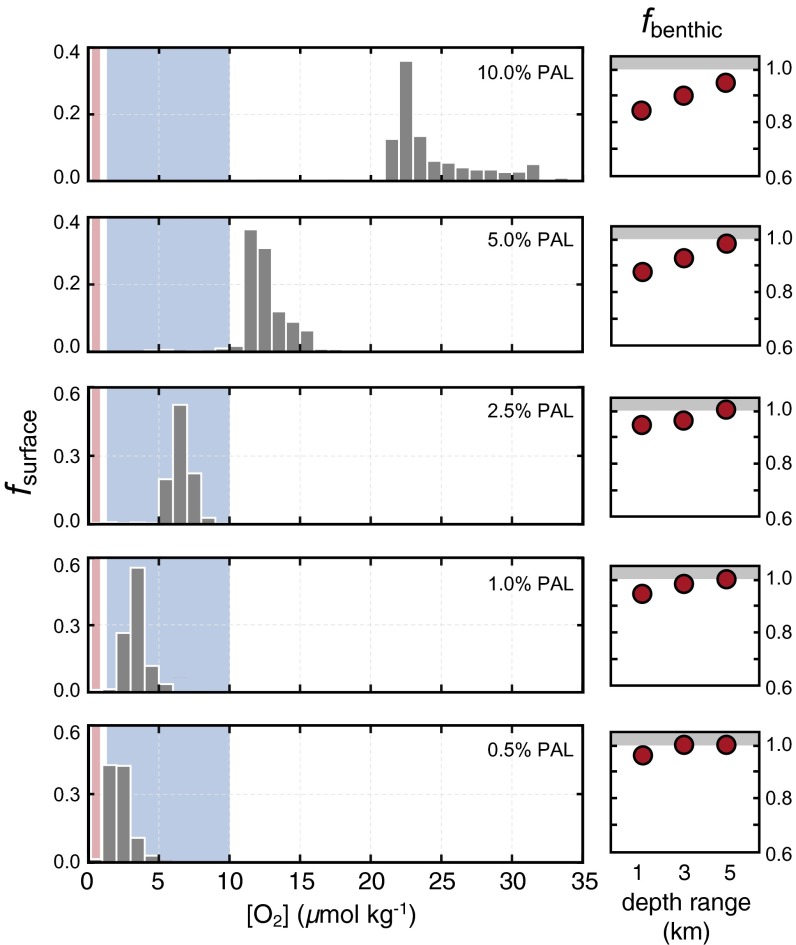
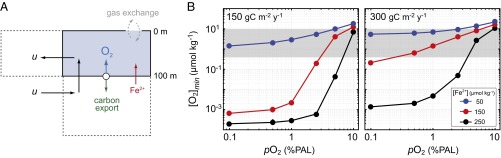
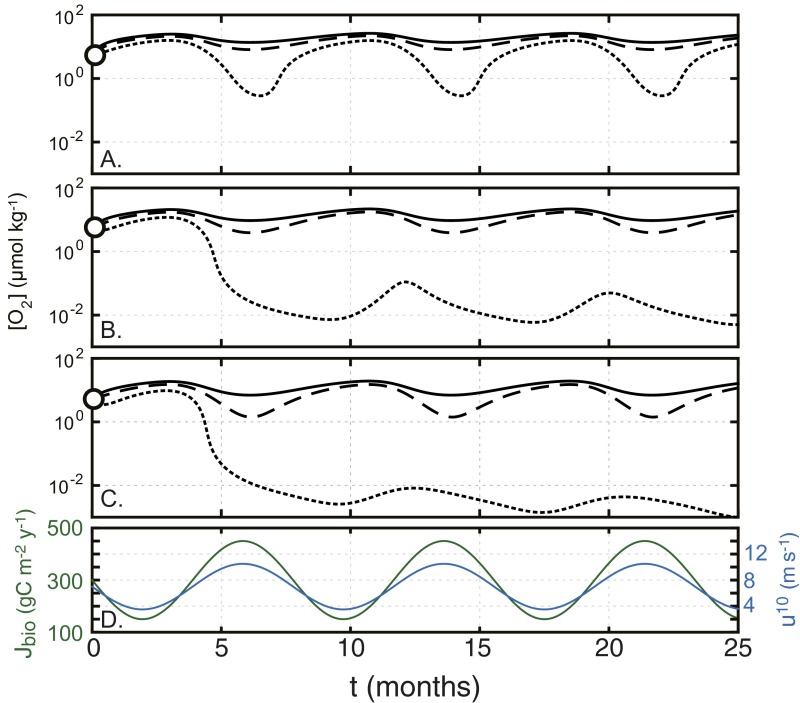
The emergence and expansion of complex eukaryotic life on Earth is linked at a basic level to the secular evolution of surface oxygen levels. However, the role that planetary redox evolution has played in controlling the timing of metazoan (animal) emergence and diversification, if any, has been intensely debated. Discussion has gravitated toward threshold levels of environmental free oxygen (O2) necessary for early evolving animals to survive under controlled conditions. However, defining such thresholds in practice is not straightforward, and environmental O2 levels can potentially constrain animal life in ways distinct from threshold O2 tolerance. Herein, we quantitatively explore one aspect of the evolutionary coupling between animal life and Earth's oxygen cycle-the influence of spatial and temporal variability in surface ocean O2 levels on the ecology of early metazoan organisms. Through the application of a series of quantitative biogeochemical models, we find that large spatiotemporal variations in surface ocean O2 levels and pervasive benthic anoxia are expected in a world with much lower atmospheric pO2 than at present, resulting in severe ecological constraints and a challenging evolutionary landscape for early metazoan life. We argue that these effects, when considered in the light of synergistic interactions with other environmental parameters and variable O2 demand throughout an organism's life history, would have resulted in long-term evolutionary and ecological inhibition of animal life on Earth for much of Middle Proterozoic time (∼1.8-0.8 billion years ago).
Keywords: Proterozoic; animals; evolution; oxygen.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Nursall JR. Oxygen as a prerequisite to the origin of the Metazoa. Nature. 1959;183(4669):1170–1172.
-
- Froelich PN, et al. Early oxidation of organic matter in pelagic sediments of the eastern equatorial Atlantic: Suboxic diagenesis. Geochim Cosmochim Acta. 1979;43(7):1075–1090.
-
- Lane N, Martin W. The energetics of genome complexity. Nature. 2010;467(7318):929–934. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
