

Chromosomal landscape of UV damage formation and repair at single-nucleotide resolution
- PMID: 27457959
- PMCID: PMC4987812
- DOI: 10.1073/pnas.1606667113
Chromosomal landscape of UV damage formation and repair at single-nucleotide resolution
Abstract
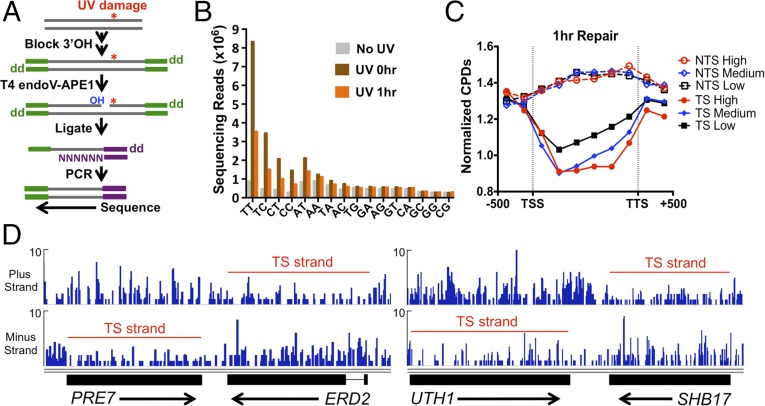
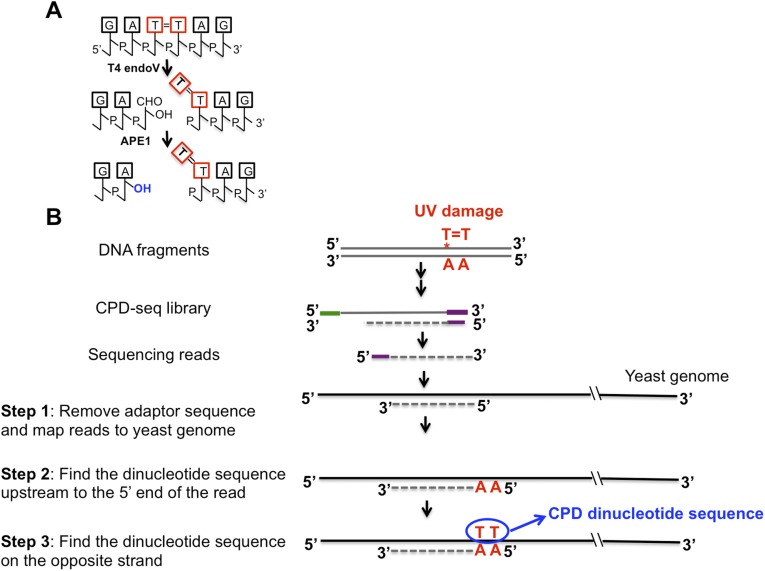
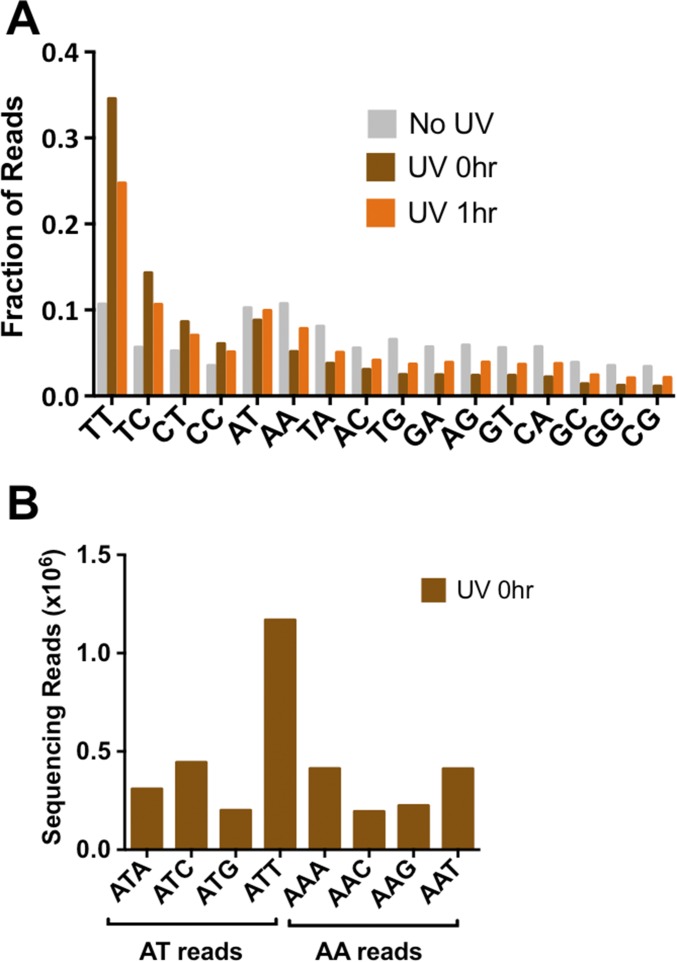
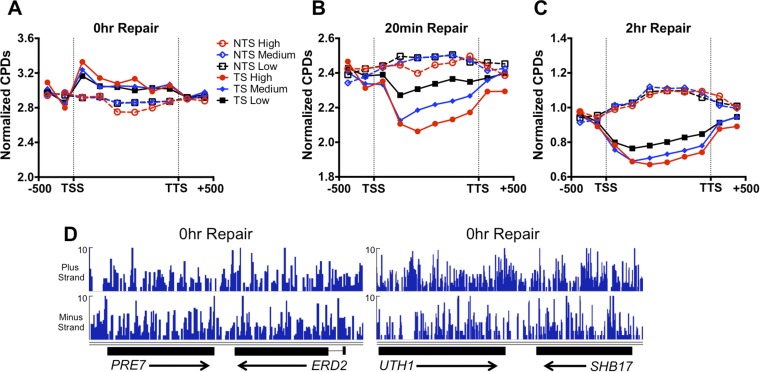
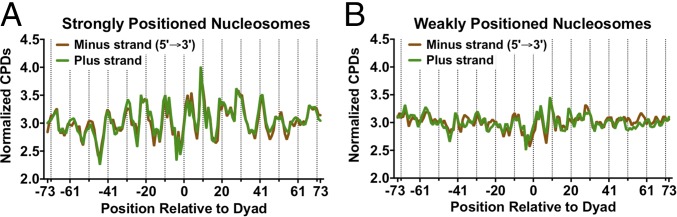
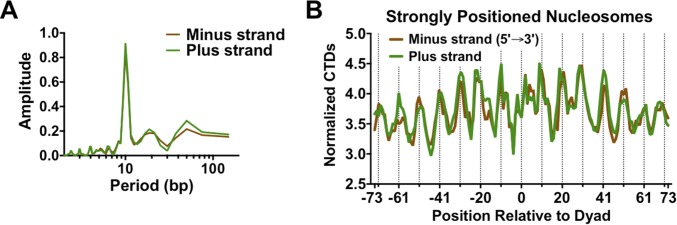
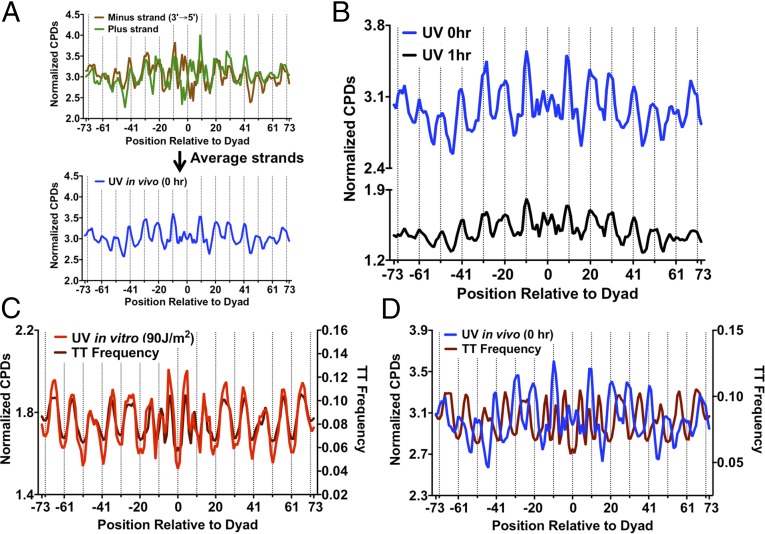
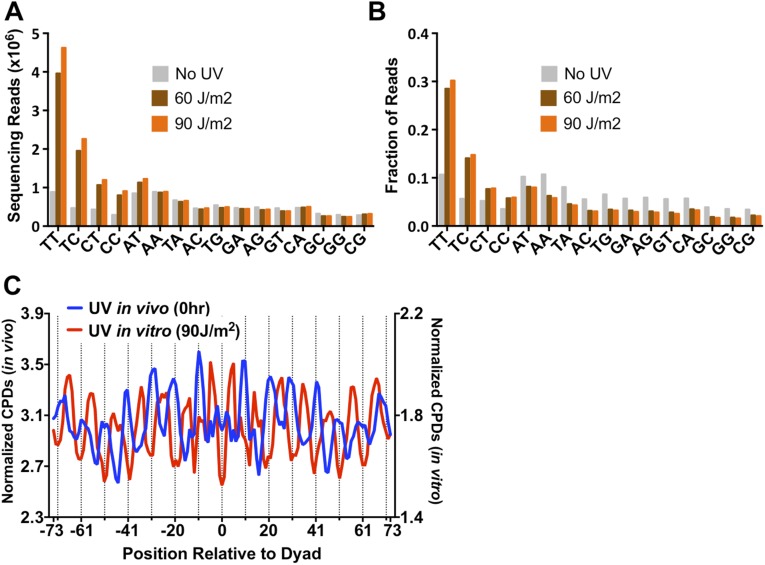
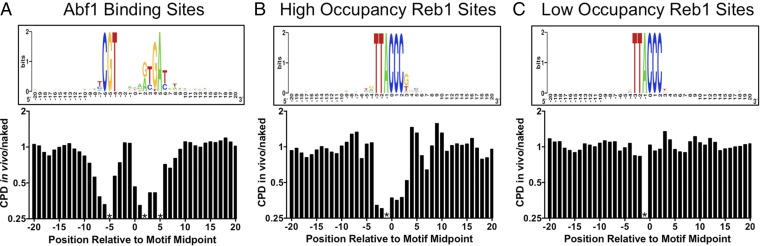
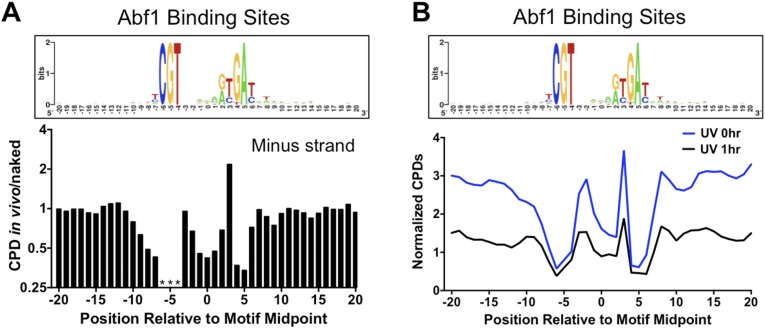
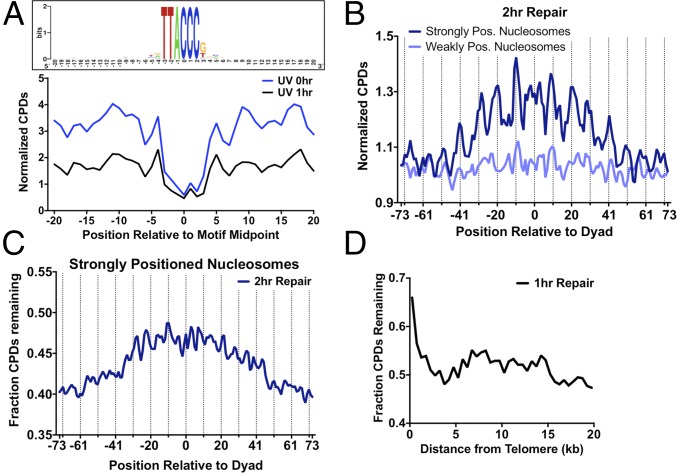
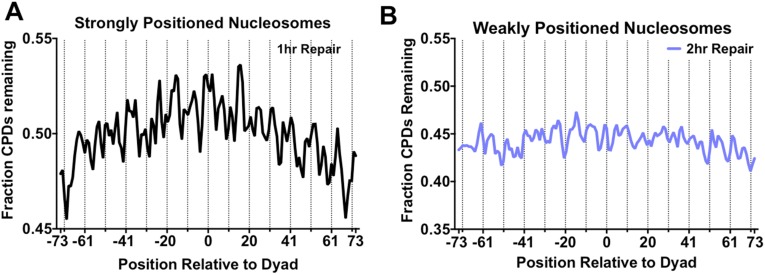
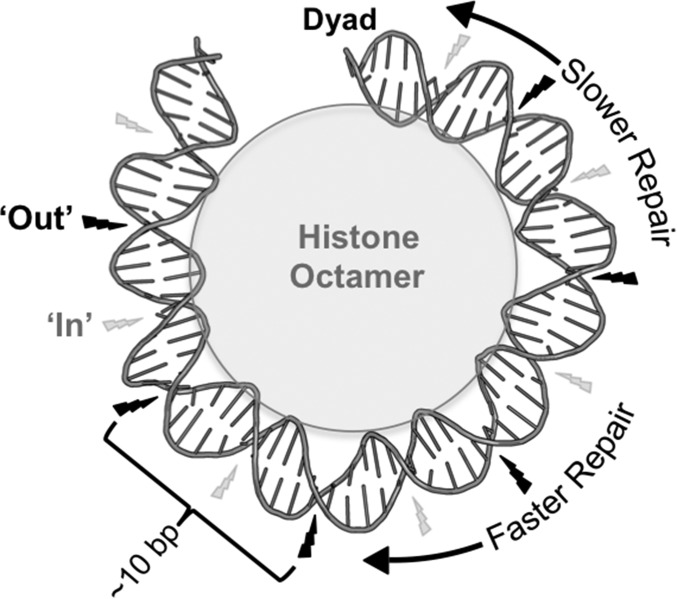
UV-induced DNA lesions are important contributors to mutagenesis and cancer, but it is not fully understood how the chromosomal landscape influences UV lesion formation and repair. Genome-wide profiling of repair activity in UV irradiated cells has revealed significant variations in repair kinetics across the genome, not only among large chromatin domains, but also at individual transcription factor binding sites. Here we report that there is also a striking but predictable variation in initial UV damage levels across a eukaryotic genome. We used a new high-throughput sequencing method, known as CPD-seq, to precisely map UV-induced cyclobutane pyrimidine dimers (CPDs) at single-nucleotide resolution throughout the yeast genome. This analysis revealed that individual nucleosomes significantly alter CPD formation, protecting nucleosomal DNA with an inward rotational setting, even though such DNA is, on average, more intrinsically prone to form CPD lesions. CPD formation is also inhibited by DNA-bound transcription factors, in effect shielding important DNA elements from UV damage. Analysis of CPD repair revealed that initial differences in CPD damage formation often persist, even at later repair time points. Furthermore, our high-resolution data demonstrate, to our knowledge for the first time, that CPD repair is significantly less efficient at translational positions near the dyad of strongly positioned nucleosomes in the yeast genome. These findings define the global roles of nucleosomes and transcription factors in both UV damage formation and repair, and have important implications for our understanding of UV-induced mutagenesis in human cancers.
Keywords: DNA damage; DNA repair; chromatin; nucleosome; transcription factor.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Friedberg EC, et al. DNA Repair and Mutagenesis. 2nd Ed ASM Press; Washington, DC: 2006.
-
- Hanawalt PC, Spivak G. Transcription-coupled DNA repair: Two decades of progress and surprises. Nat Rev Mol Cell Biol. 2008;9(12):958–970. - PubMed
-
- Marteijn JA, Lans H, Vermeulen W, Hoeijmakers JH. Understanding nucleotide excision repair and its roles in cancer and ageing. Nat Rev Mol Cell Biol. 2014;15(7):465–481. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
