

Age-related changes in tonic activation of presynaptic versus extrasynaptic γ-amniobutyric acid type B receptors in rat medial prefrontal cortex
- PMID: 27459929
- PMCID: PMC5235322
- DOI: 10.1016/j.neurobiolaging.2016.05.015
Age-related changes in tonic activation of presynaptic versus extrasynaptic γ-amniobutyric acid type B receptors in rat medial prefrontal cortex
Abstract
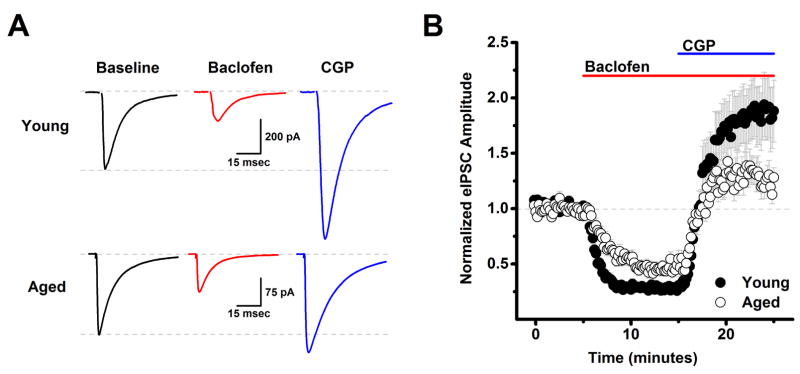
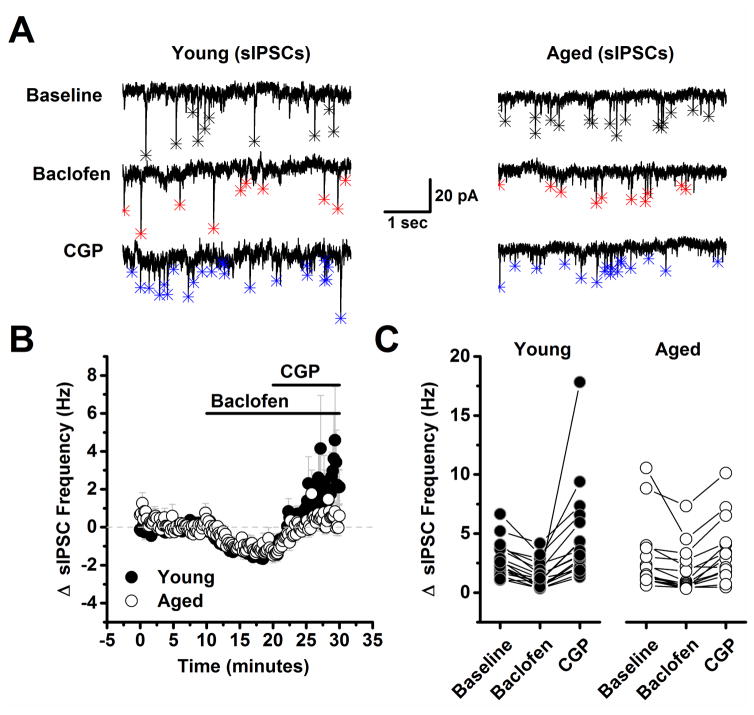
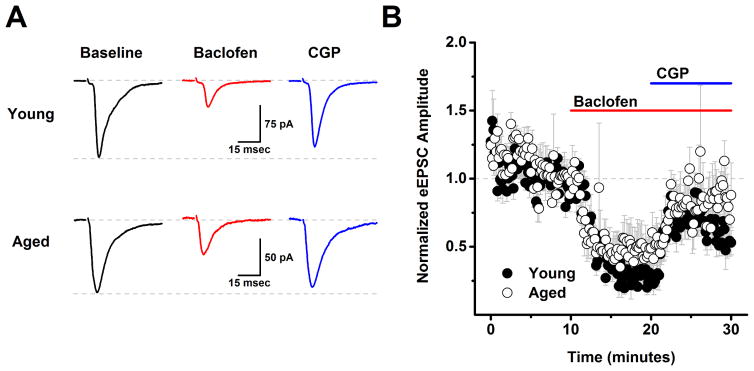
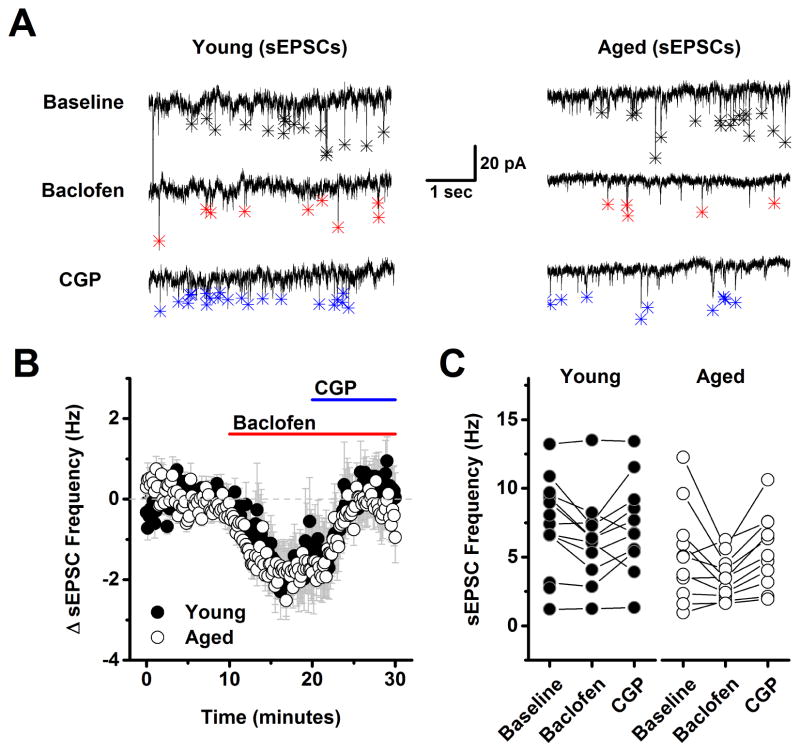
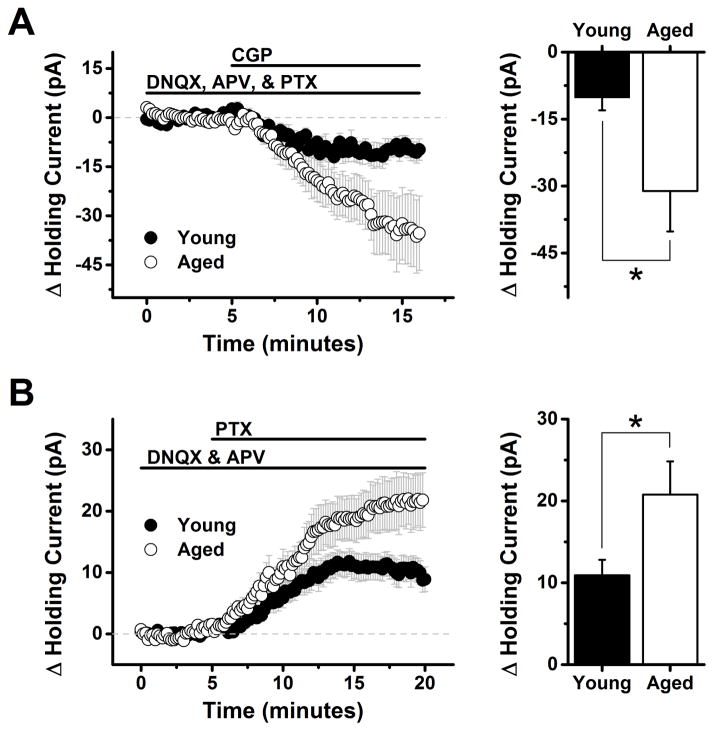
The present study examined the effect of age on both glutamatergic and γ-aminobutyric acid mediated (GABAergic) signaling in the rodent medial prefrontal cortex (mPFC), with an emphasis on revealing novel changes contributing to increased inhibition in age. Whole-cell patch clamp recordings were obtained from layer 2/3 mPFC pyramidal neurons in acute cortical slices prepared from either young (4 months) or aged (20-24 months) male F344 rats. Results indicated that GABAB receptors on GABAergic, but not on glutamatergic, inputs to layer 2/3 pyramidal cells are tonically activated by ambient GABA in young animals and further demonstrated that this form of tonic inhibition is significantly attenuated in aged mPFC. Moreover, concurrent with loss of tonic presynaptic GABAB autoreceptor activation, layer 2/3 pyramidal cells in aged mPFC are subjected to increased tonic activation of extrasynaptic GABAA and GABAB receptors. These data demonstrate a shift in the site of GABAB receptor-mediated inhibitory tone in the aged mPFC that clearly promotes increased inhibition of pyramidal cells in aged animals, and that may plausibly contribute to impaired executive function.
Keywords: Age-related cognitive decline; GABA(A); GABA(B); L2/3 pyramidal neurons; Prefrontal cortex; Working memory.
Copyright © 2016 Elsevier Inc. All rights reserved.
Conflict of interest statement
statement The authors have no conflicts of interest, and all animal procedures were performed in compliance with the Institutional Animal Care and Use Committee (IACUC) of the University of Florida.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
