

Internalized Tau sensitizes cells to stress by promoting formation and stability of stress granules
- PMID: 27460788
- PMCID: PMC4962319
- DOI: 10.1038/srep30498
Internalized Tau sensitizes cells to stress by promoting formation and stability of stress granules
Abstract
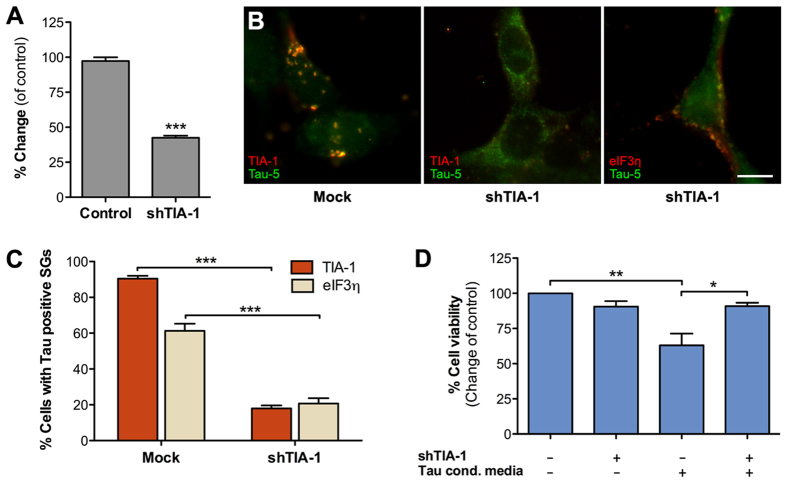
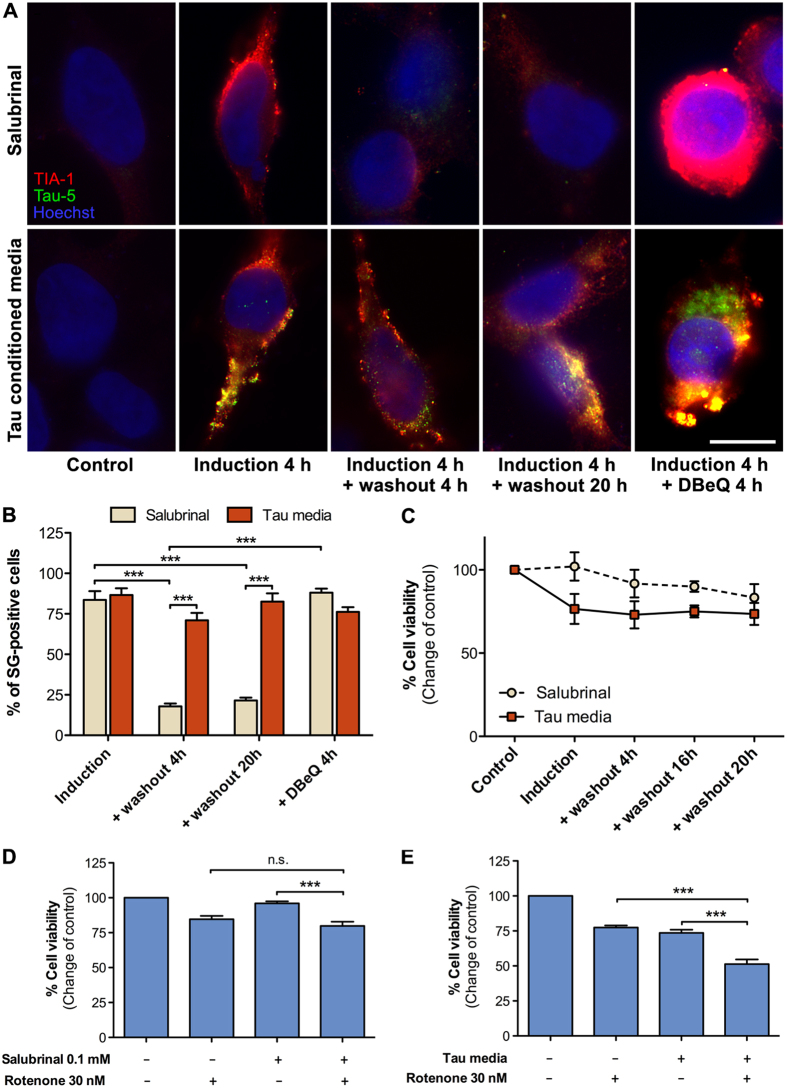
Stress granules are membrane-less RNA- and RNA-binding protein-containing complexes that are transiently assembled in stressful conditions to promote cell survival. Several stress granule-associated RNA-binding proteins have been associated with neurodegenerative diseases. In addition, a close link was recently identified between the stress granule core-nucleating protein TIA-1 and Tau. Tau is a central pathological protein in Alzheimer's disease and other tauopathies, and misfolded, aggregated Tau is capable of propagating pathology via cell-to-cell transmission. Here we show that following internalization hyperphosphorylated extracellular Tau associates with stress granules in a TIA-1 dependent manner. Cytosolic Tau normally only weakly interacts with TIA-1 but mutations mimicking abnormal phosphorylation promote this interaction. We show that internalized Tau significantly delays normal clearance of stress granules in the recipient cells sensitizing them to secondary stress. These results suggest that secreted Tau species may have properties, likely related to its hyperphosphorylation and oligomerization, which promote pathological association of internalized Tau with stress granules altering their dynamics and reducing cell viability. We suggest that stress granules and TIA-1 play a central role in the cell-to-cell transmission of Tau pathology.
Conflict of interest statement
H.J.H. is a co-founder and shareholder of Herantis Pharma plc, which is not related to this study. The other authors declare no conflict of interests.
Figures
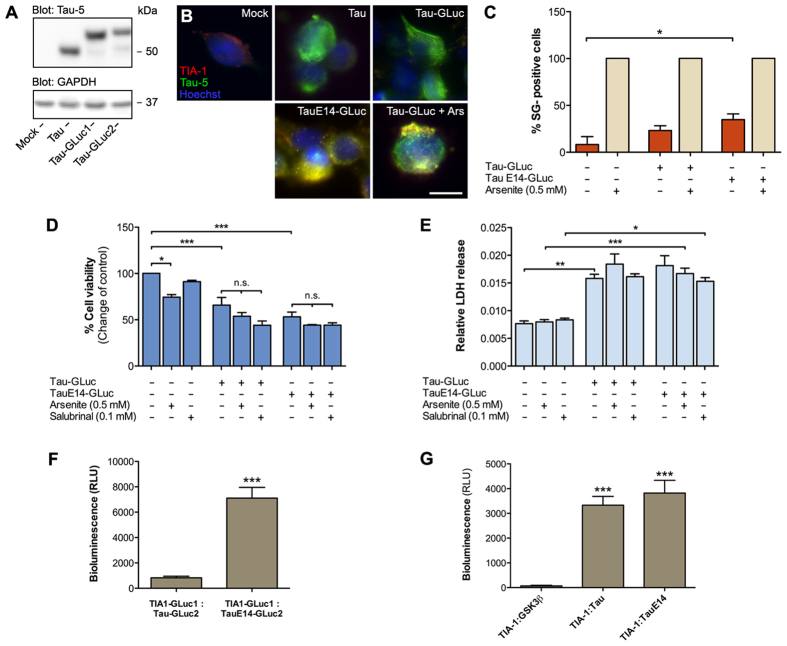
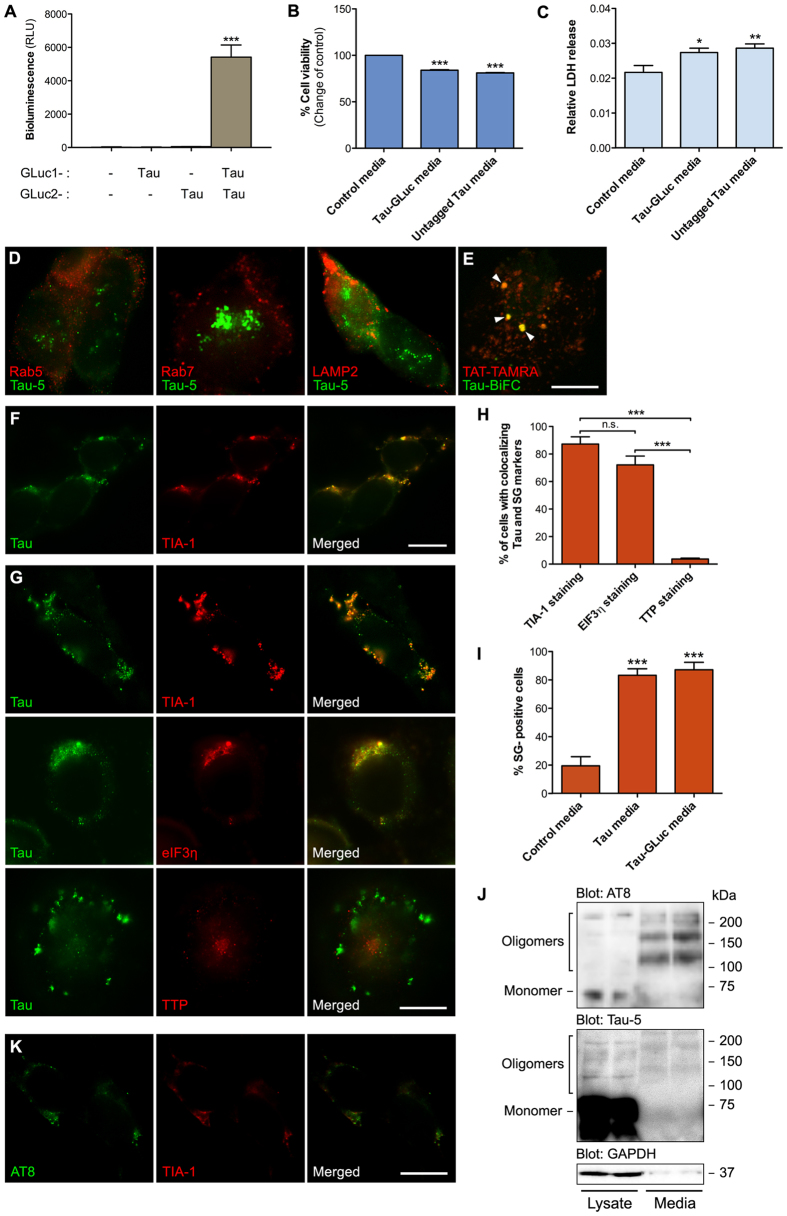
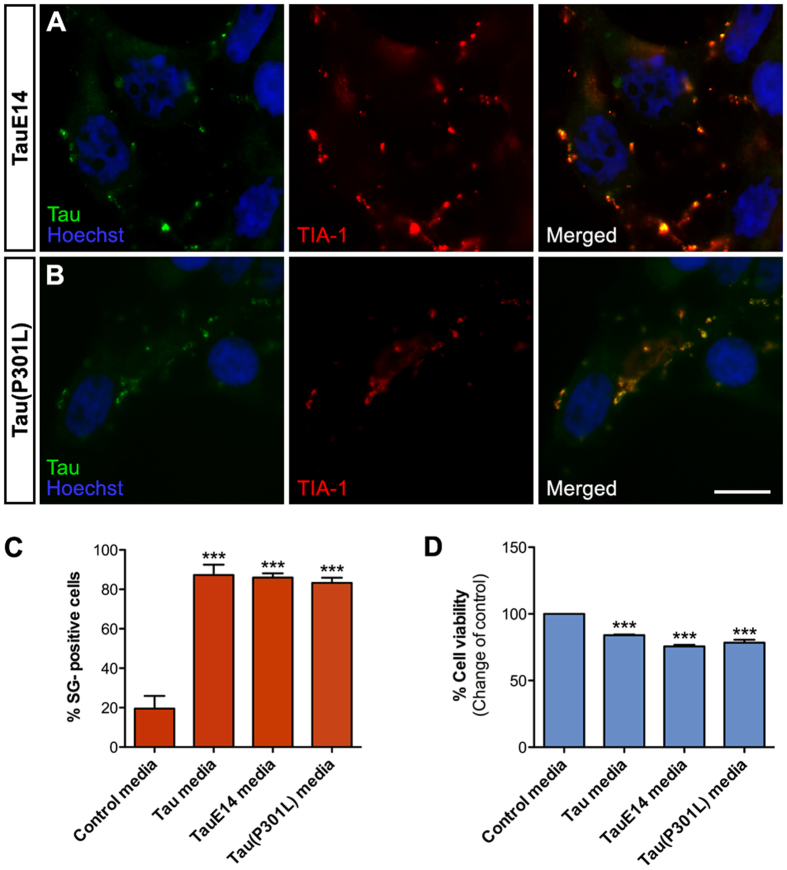
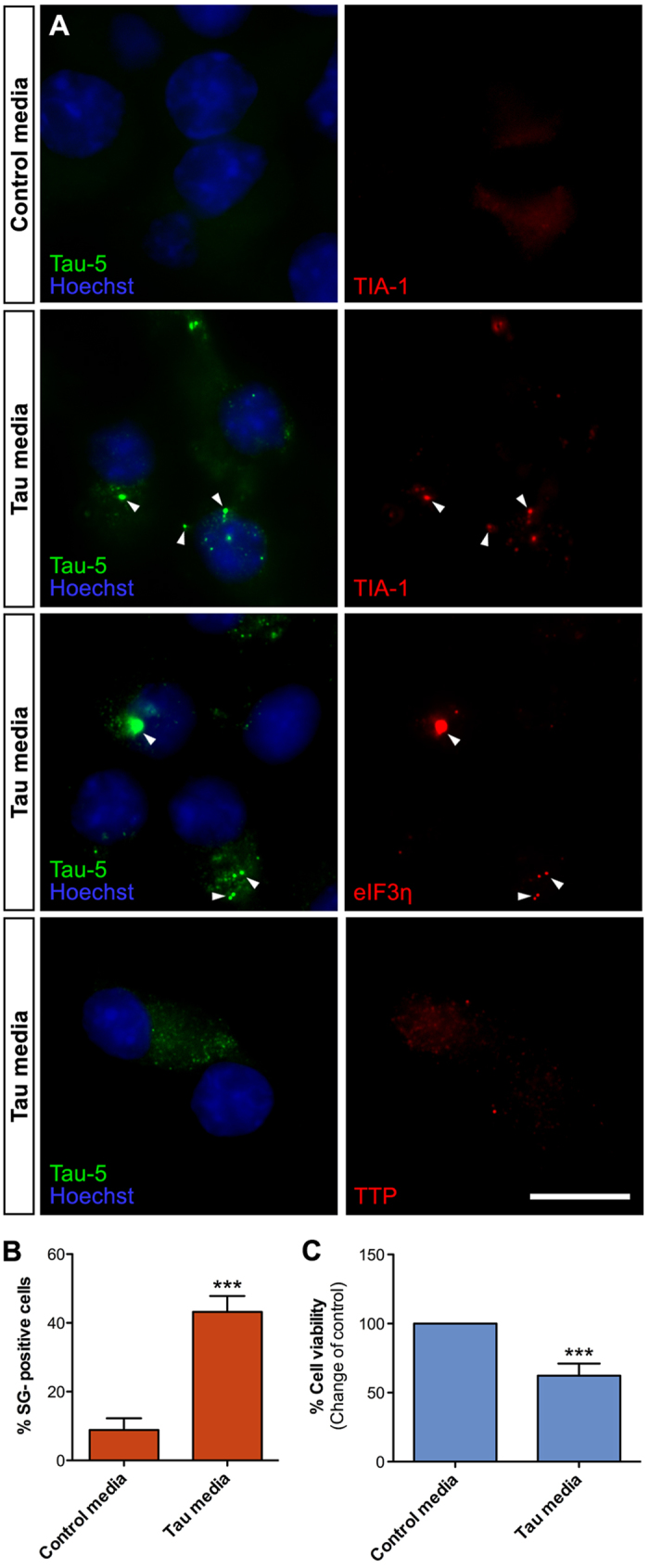
References
-
- Neelamraju Y., Hashemikhabir S. & Janga S. C. The human RBPome: from genes and proteins to human disease. J Proteomics. 127, 61–70 (2015). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
