

Differentiation of distinct long-lived memory CD4 T cells in intestinal tissues after oral Listeria monocytogenes infection
- PMID: 27461178
- PMCID: PMC5272904
- DOI: 10.1038/mi.2016.66
Differentiation of distinct long-lived memory CD4 T cells in intestinal tissues after oral Listeria monocytogenes infection
Abstract
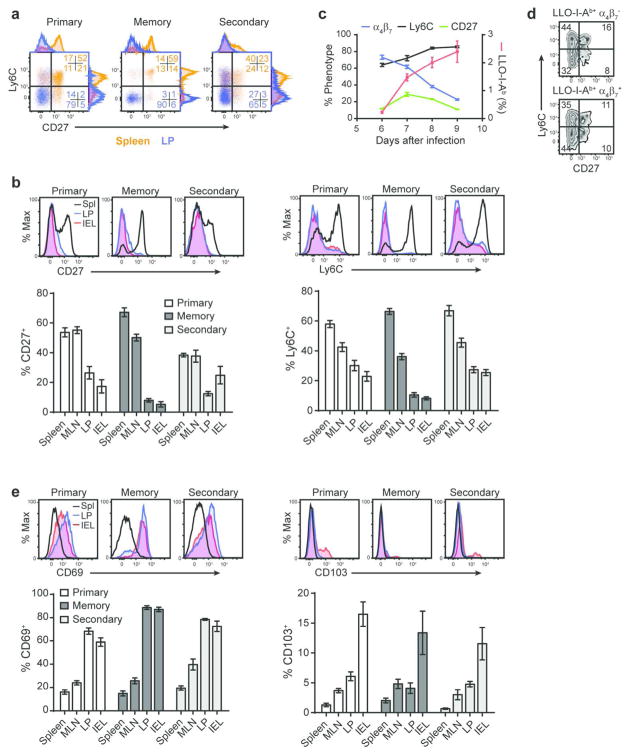
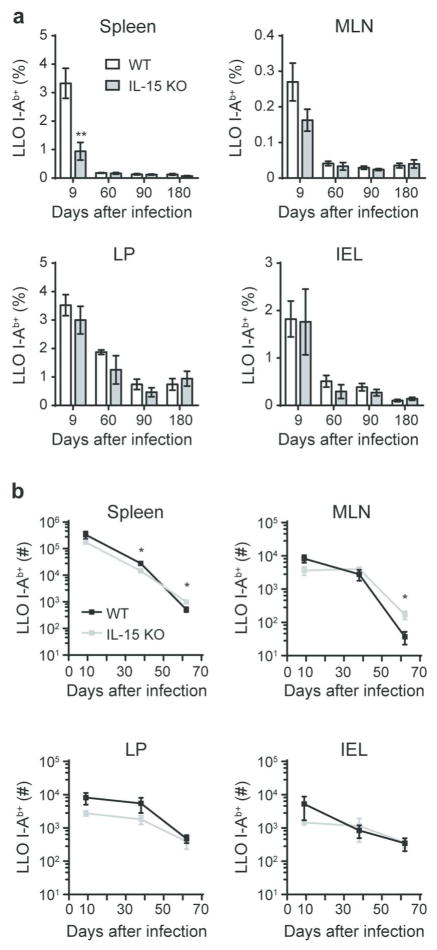
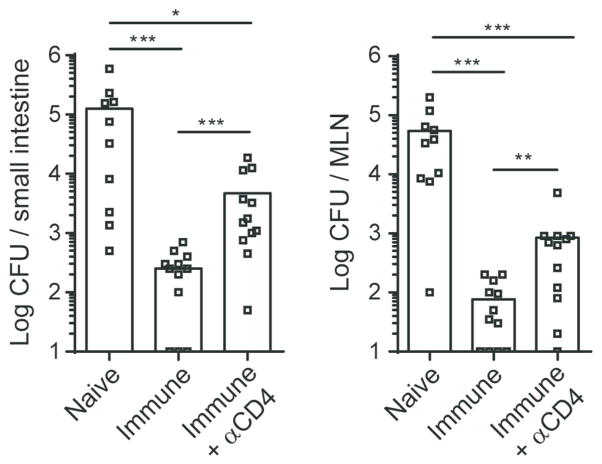
Mucosal antigen-specific CD4 T-cell responses to intestinal pathogens remain incompletely understood. Here we examined the CD4 T-cell response after oral infection with an internalin A 'murinized' Listeria monocytogenes (Lm). Oral Lm infection induced a robust endogenous listeriolysin O (LLO)-specific CD4 T-cell response with distinct phenotypic and functional characteristics in the intestine. Circulating LLO-specific CD4 T cells transiently expressed the 'gut-homing' integrin α4β7 and accumulated in the intestinal lamina propria and epithelium where they were maintained independent of interleukin (IL)-15. The majority of intestinal LLO-specific CD4 T cells were CD27- Ly6C- and CD69+ CD103- while the lymphoid LLO-specific CD4 T cells were heterogeneous based on CD27 and Ly6C expression and predominately CD69-. LLO-specific effector CD4 T cells transitioned into a long-lived memory population that phenotypically resembled their parent effectors and displayed hallmarks of residency. In addition, intestinal effector and memory CD4 T cells showed a predominant polyfunctional Th1 profile producing IFNγ, TNFα, and IL-2 at high levels with minimal but detectable levels of IL-17A. Depletion of CD4 T cells in immunized mice led to elevated bacterial burden after challenge infection highlighting a critical role for memory CD4 T cells in controlling intestinal intracellular pathogens.
Conflict of interest statement
Figures


Similar articles
-
Cytolysin-dependent escape of the bacterium from the phagosome is required but not sufficient for induction of the Th1 immune response against Listeria monocytogenes infection: distinct role of Listeriolysin O determined by cytolysin gene replacement.Infect Immun. 2007 Aug;75(8):3791-801. doi: 10.1128/IAI.01779-06. Epub 2007 May 21. Infect Immun. 2007. PMID: 17517863 Free PMC article.
-
Organ-specific CD4+ T cell response during Listeria monocytogenes infection.J Immunol. 2002 Jun 15;168(12):6382-7. doi: 10.4049/jimmunol.168.12.6382. J Immunol. 2002. PMID: 12055256
-
A gold glyco-nanoparticle carrying a Listeriolysin O peptide and formulated with Advax™ delta inulin adjuvant induces robust T-cell protection against listeria infection.Vaccine. 2015 Mar 17;33(12):1465-73. doi: 10.1016/j.vaccine.2015.01.062. Epub 2015 Feb 7. Vaccine. 2015. PMID: 25659269
-
Intestinal and splenic T cell responses to enteric Listeria monocytogenes infection: distinct repertoires of responding CD8 T lymphocytes.J Immunol. 2001 Mar 15;166(6):4065-73. doi: 10.4049/jimmunol.166.6.4065. J Immunol. 2001. PMID: 11238655
-
Mechanisms and immunological effects of apoptosis caused by Listeria monocytogenes.Adv Immunol. 2012;113:157-74. doi: 10.1016/B978-0-12-394590-7.00001-4. Adv Immunol. 2012. PMID: 22244583 Review.
Cited by
-
T Cell Receptor Is Required for Differentiation, but Not Maintenance, of Intestinal CD4+ Intraepithelial Lymphocytes.Immunity. 2020 Nov 17;53(5):1001-1014.e20. doi: 10.1016/j.immuni.2020.09.003. Epub 2020 Oct 5. Immunity. 2020. PMID: 33022229 Free PMC article.
-
Divergence of Tissue-Memory T Cells: Distribution and Function-Based Classification.Cold Spring Harb Perspect Biol. 2020 Oct 1;12(10):a037762. doi: 10.1101/cshperspect.a037762. Cold Spring Harb Perspect Biol. 2020. PMID: 32816841 Free PMC article. Review.
-
Regulation of Th17 cells by P. UF1 against systemic Listeria monocytogenes infection.Gut Microbes. 2018;9(3):279-287. doi: 10.1080/19490976.2017.1417731. Epub 2018 Feb 8. Gut Microbes. 2018. PMID: 29420115 Free PMC article.
-
T lymphocytes in the intestinal mucosa: defense and tolerance.Cell Mol Immunol. 2019 Mar;16(3):216-224. doi: 10.1038/s41423-019-0208-2. Epub 2019 Feb 20. Cell Mol Immunol. 2019. PMID: 30787416 Free PMC article. Review.
-
CD4+ Tissue-resident Memory T Cells Expand and Are a Major Source of Mucosal Tumour Necrosis Factor α in Active Crohn's Disease.J Crohns Colitis. 2019 Jul 25;13(7):905-915. doi: 10.1093/ecco-jcc/jjz010. J Crohns Colitis. 2019. PMID: 30715262 Free PMC article.
References
-
- Harrington LE, et al. Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nat Immunol. 2005;6:1123–1132. - PubMed
-
- Becattini S, et al. T cell immunity. Functional heterogeneity of human memory CD4(+) T cell clones primed by pathogens or vaccines. Science. 2015;347:400–406. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
