

Targeting the γ-/β-secretase interaction reduces β-amyloid generation and ameliorates Alzheimer's disease-related pathogenesis
- PMID: 27462420
- PMCID: PMC4860824
- DOI: 10.1038/celldisc.2015.21
Targeting the γ-/β-secretase interaction reduces β-amyloid generation and ameliorates Alzheimer's disease-related pathogenesis
Abstract
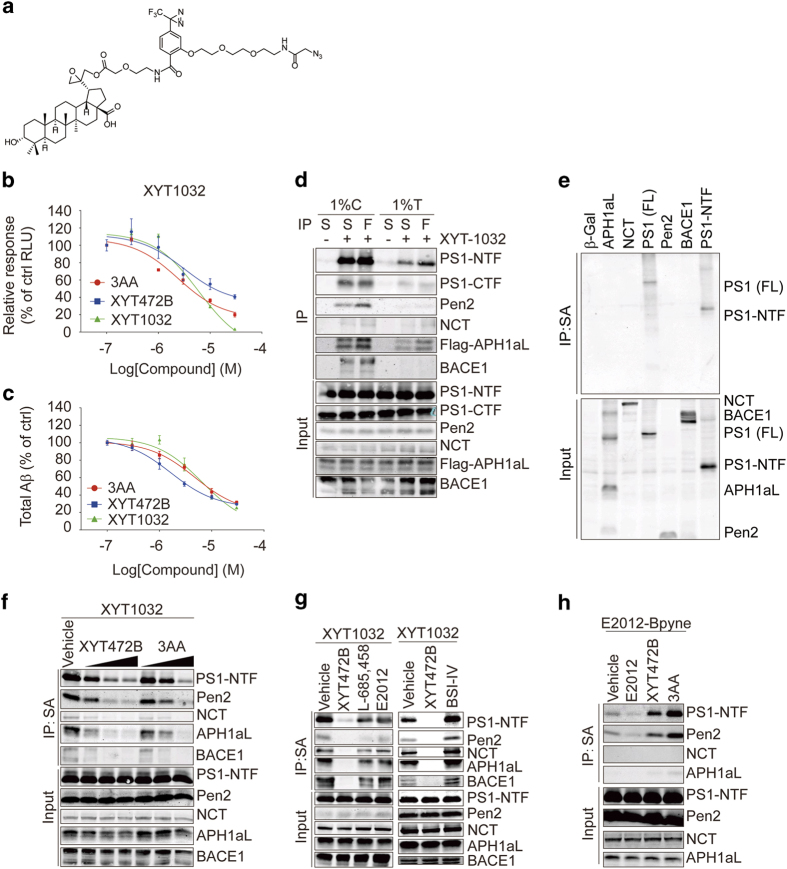
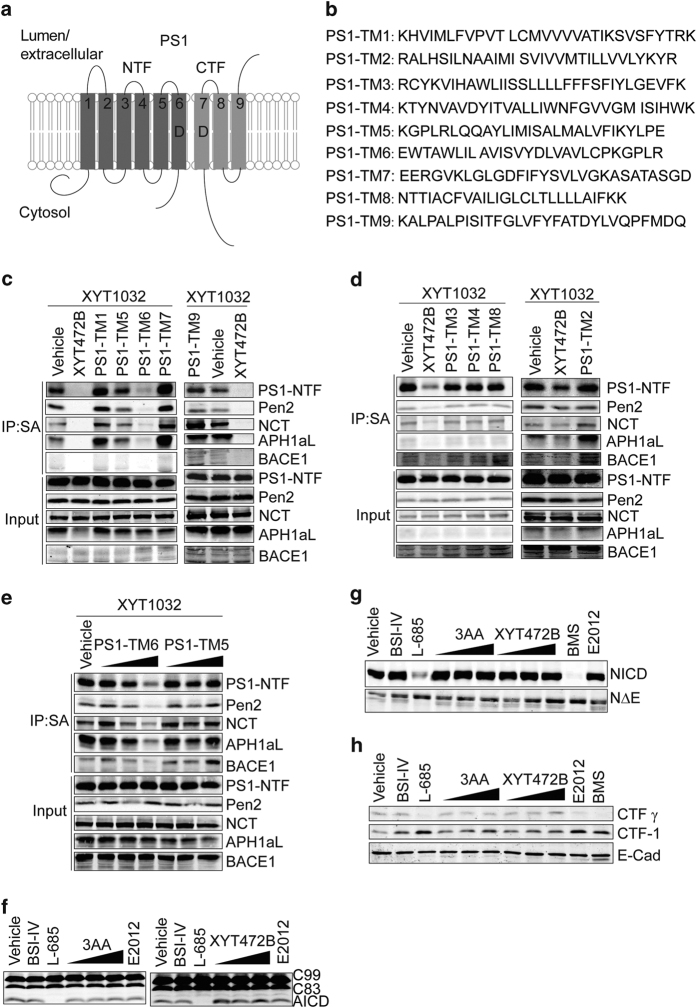
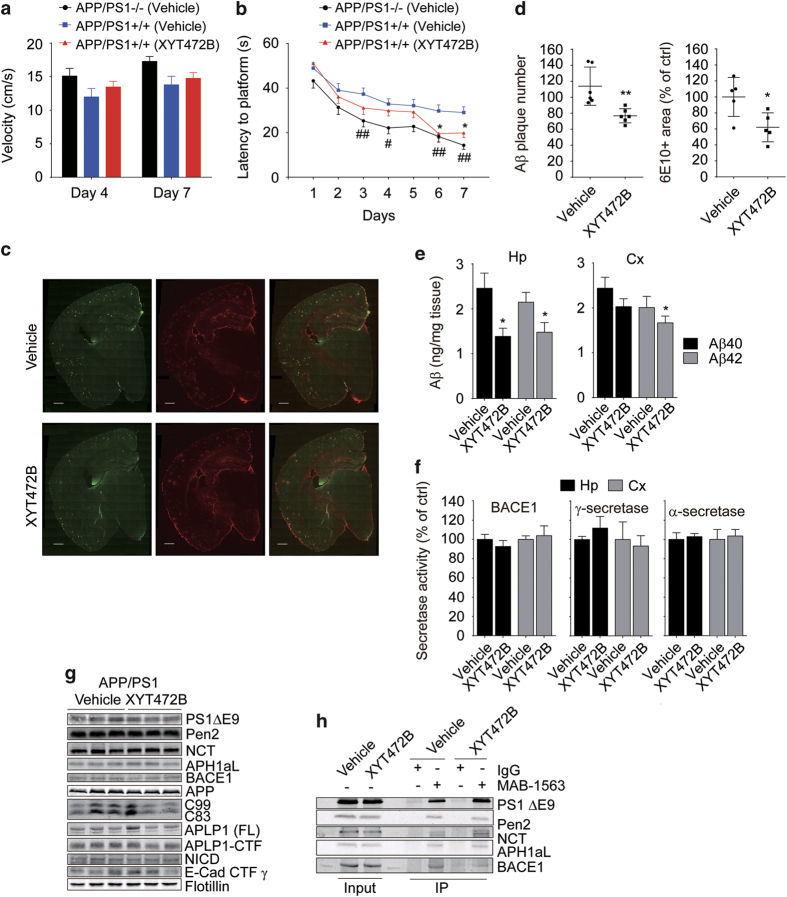
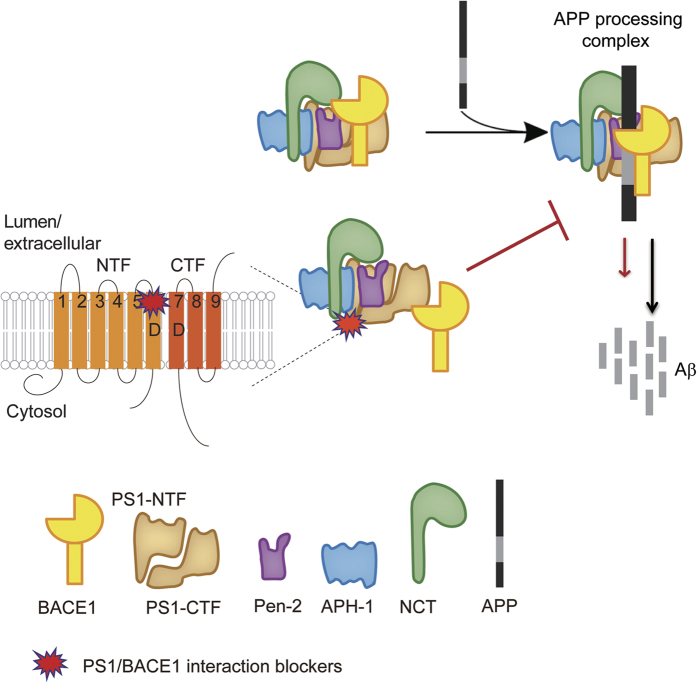
Despite decades of intense global effort, no disease-modifying drugs for Alzheimer's disease have emerged. Molecules targeting catalytic activities of γ-secretase or β-site APP-cleaving enzyme 1 (BACE1) have been beset by undesired side effects. We hypothesized that blocking the interaction between BACE1 and γ-secretase subunit presenilin-1 (PS1) might offer an alternative strategy to selectively suppress Aβ generation. Through high-throughput screening, we discovered that 3-α-akebonoic acid (3AA) interferes with PS1/BACE1 interaction and reduces Aβ production. Structural analogs of 3AA were systematically synthesized and the functional analog XYT472B was identified. Photo-activated crosslinking and biochemical competition assays showed that 3AA and XYT472B bind to PS1, interfere with PS1/BACE1 interaction, and reduce Aβ production, whereas sparing secretase activities. Furthermore, treatment of APP/PS1 mice with XYT472B alleviated cognitive dysfunction and Aβ-related pathology. Together, our results indicate that chemical interference of PS1/BACE1 interaction is a promising strategy for Alzheimer's disease therapeutics.
Keywords: Alzheimer’s disease; Aβ generation; PS1/BACE1 interaction.
Figures
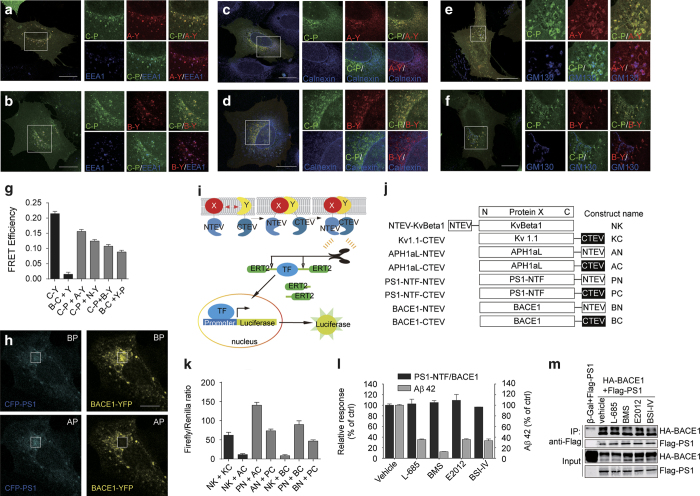
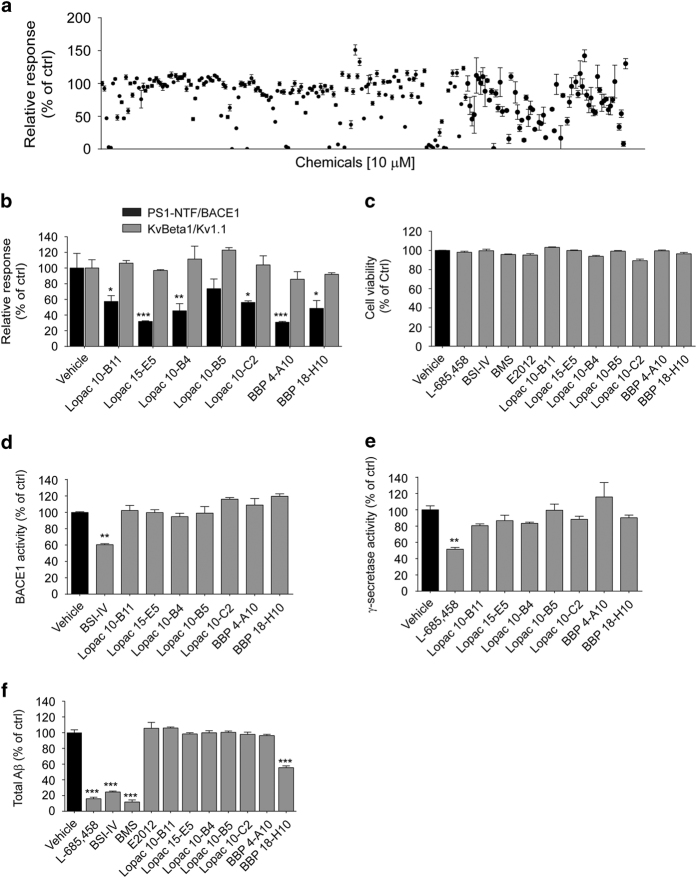
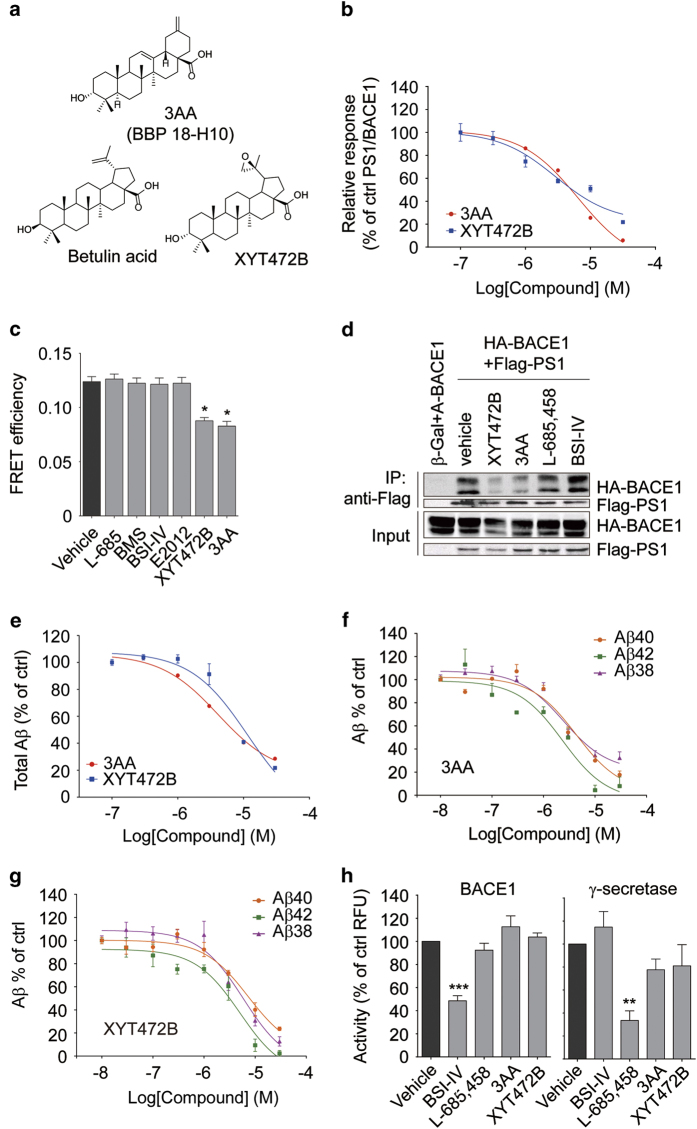
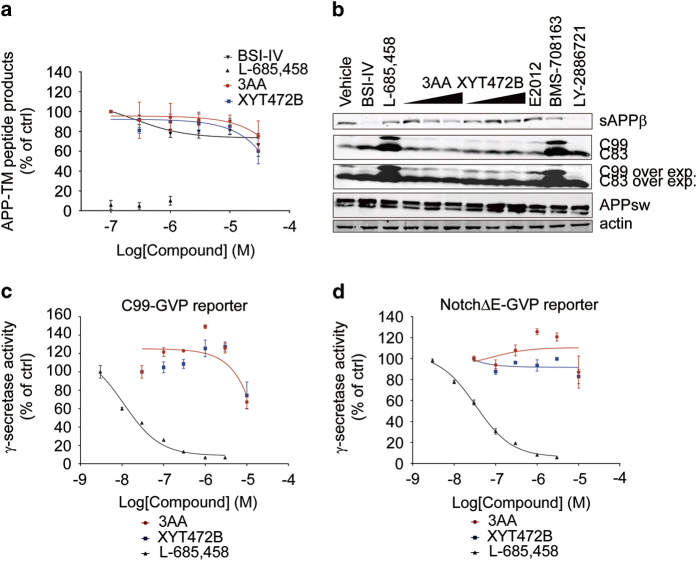
References
-
- Goedert M , Spillantini MG . A century of Alzheimer's disease. Science 2006; 314: 777–781. - PubMed
-
- Hardy J , Selkoe DJ . The amyloid hypothesis of Alzheimer's disease: progress and problems on the road to therapeutics. Science 2002; 297: 353–356. - PubMed
-
- Karran E , Mercken M , De Strooper B . The amyloid cascade hypothesis for Alzheimer's disease: an appraisal for the development of therapeutics. Nat Rev Drug Discov 2011; 10: 698–712. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
