

Effects of Aging and Oxidative Stress on Spermatozoa of Superoxide-Dismutase 1- and Catalase-Null Mice
- PMID: 27465136
- PMCID: PMC5333935
- DOI: 10.1095/biolreprod.116.141671
Effects of Aging and Oxidative Stress on Spermatozoa of Superoxide-Dismutase 1- and Catalase-Null Mice
Abstract
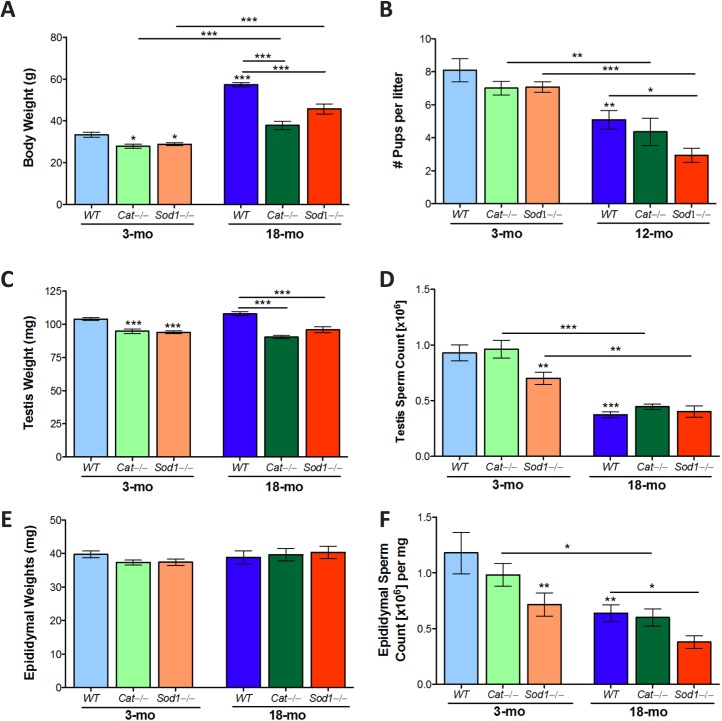
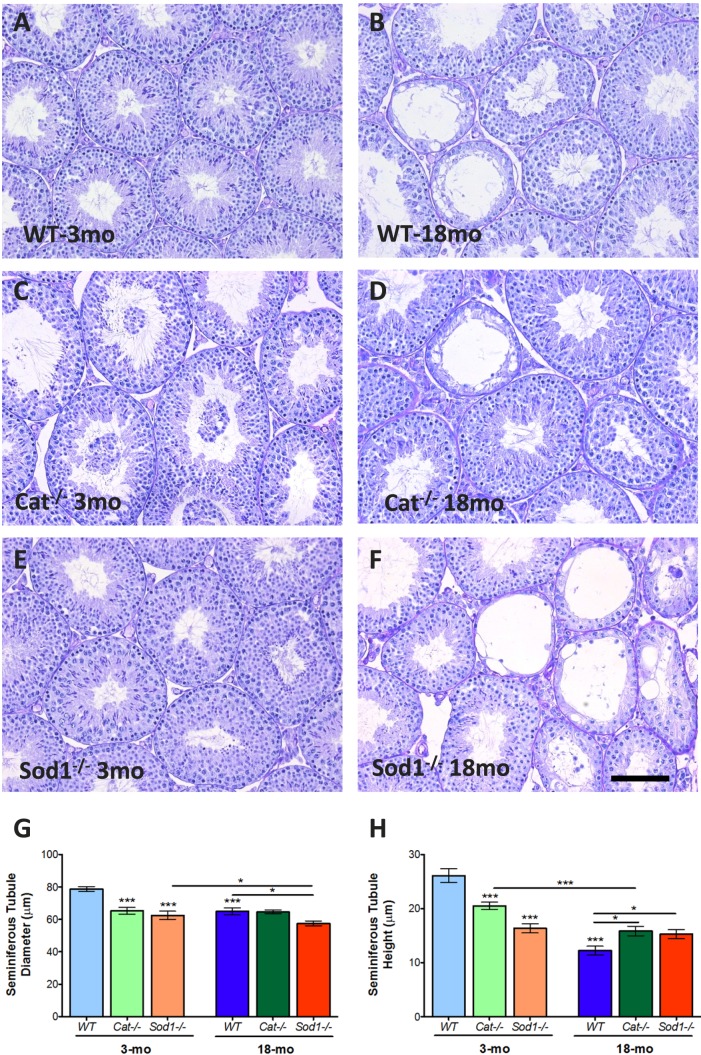
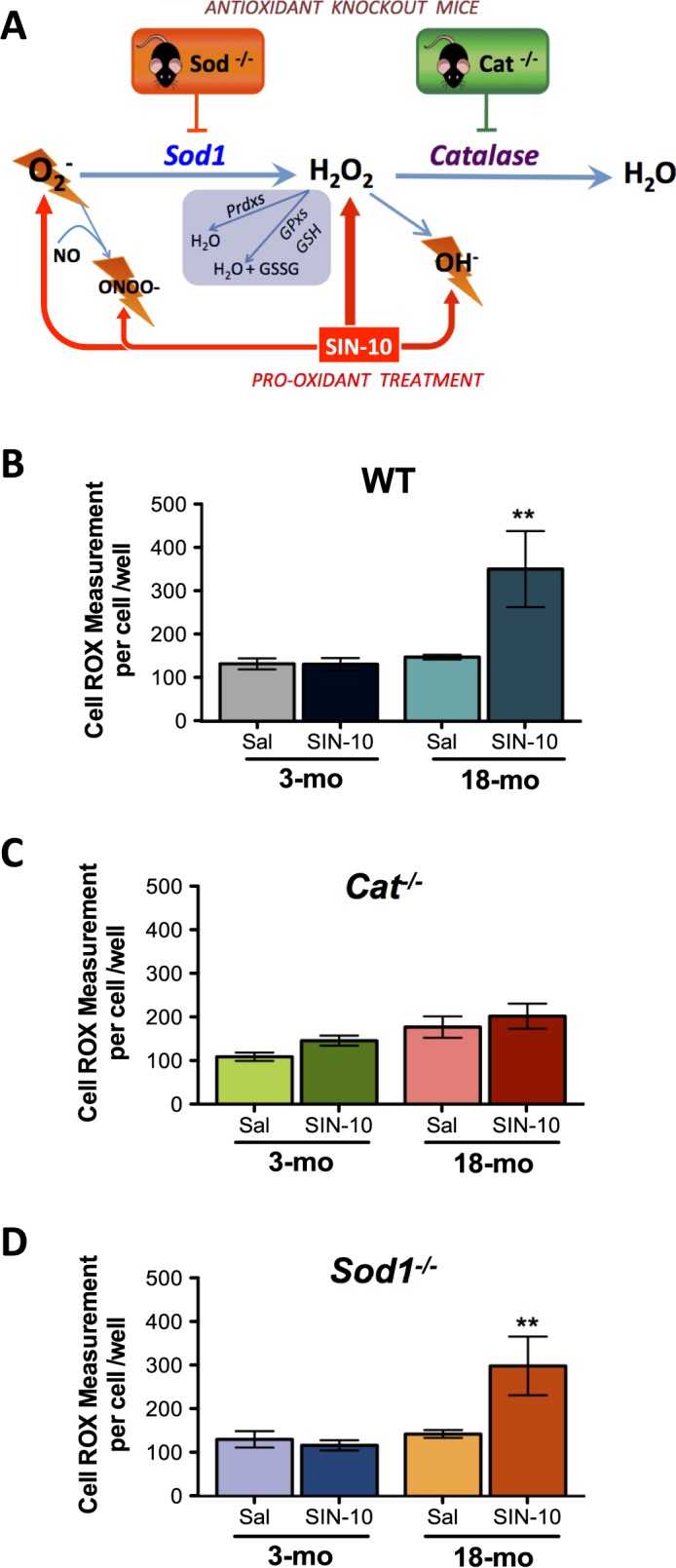
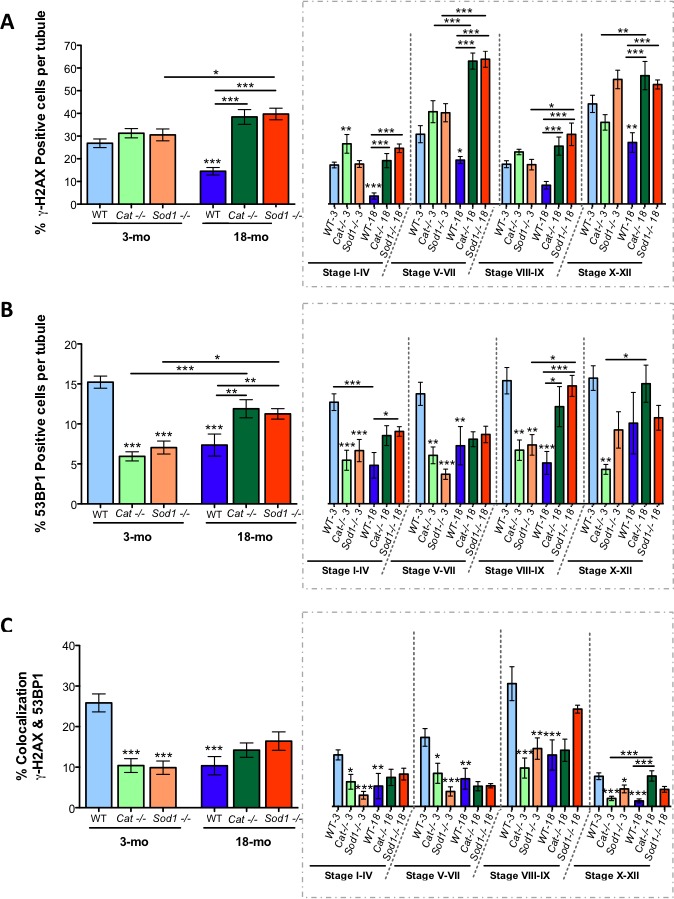
Advanced paternal age is linked to complications in pregnancy and genetic diseases in offspring. Aging results in excess reactive oxygen species (ROS) and DNA damage in spermatozoa; this damage can be transmitted to progeny with detrimental consequences. Although there is a loss of antioxidants with aging, the impact on aging male germ cells of the complete absence of either catalase (CAT) or superoxide dismutase 1 (SOD1) has not been investigated. We used CAT-null (Cat(-/-)) and SOD1-null (Sod(-/-)) mice to determine whether loss of these antioxidants increases germ cell susceptibility to redox dysfunction with aging. Aging reduced fertility and the numbers of Sertoli and germ cells in all mice. Aged Sod(-/-) mice displayed an increased loss of fertility compared to aged wild-type mice. Treatment with the pro-oxidant SIN-10 increased ROS in spermatocytes of aged wild-type and Sod(-/-) mice, while aged Cat(-/-) mice were able to neutralize this ROS. The antioxidant peroxiredoxin 1 (PRDX1) increased with age in wild-type and Cat(-/-) mice but was consistently low in young and aged Sod(-/-) mice. DNA damage and repair markers (γ-H2AX and 53BP1) were reduced with aging and lower in young Sod(-/-) and Cat(-/-) mice. Colocalization of γ-H2AX and 53BP1 suggested active repair in young wild-type mice but reduced in young Cat(-/-) and in Sod(-/-) mice and with age. Oxidative DNA damage (8-oxodG) increased in young Sod(-/-) mice and with age in all mice. These studies show that aged Sod(-/-) mice display severe redox dysfunction, while wild-type and Cat(-/-) mice have compensatory mechanisms to partially alleviate oxidative stress and reduce age-related DNA damage in spermatozoa. Thus, SOD1 but not CAT is critical to the maintenance of germ cell quality with aging.
Keywords: Sertoli cells; aging; oxidative stress; spermatogenesis.
© 2016 by the Society for the Study of Reproduction, Inc.
Figures



References
-
- Lawson G, Fletcher R. Delayed fatherhood. J Fam Plan Reprod Health Care. 2014;40:283–288. - PubMed
-
- Hassan MA, Killick SR. Effect of male age on fertility: evidence for the decline in male fertility with increasing age. Fertil Steril. 2003;79(suppl 3):1520–1527. - PubMed
-
- Serre V, Robaire B. Paternal age affects fertility and progeny outcome in the Brown Norway rat. Fertil Steril. 1998;70:625–631. - PubMed
-
- Kleinhaus K, Perrin M, Friedlander Y, Paltiel O, Malaspina D, Harlap S. Paternal age and spontaneous abortion. Obstet Gynecol. 2006;108:369–377. - PubMed
-
- Martin AK, Robinson G, Reutens D, Mowry B. Clinical and parental age characteristics of rare copy number variant burden in patients with schizophrenia. Am J Med Genet B Neuropsychiatr Genet. 2015;168B:374–382. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
