

Sensory hypo-excitability in a rat model of fetal development in Fragile X Syndrome
- PMID: 27465362
- PMCID: PMC4964352
- DOI: 10.1038/srep30769
Sensory hypo-excitability in a rat model of fetal development in Fragile X Syndrome
Erratum in
-
Erratum: Sensory hypo-excitability in a rat model of fetal development in Fragile X Syndrome.Sci Rep. 2017 Mar 15;7:44515. doi: 10.1038/srep44515. Sci Rep. 2017. PMID: 28294163 Free PMC article. No abstract available.
Abstract
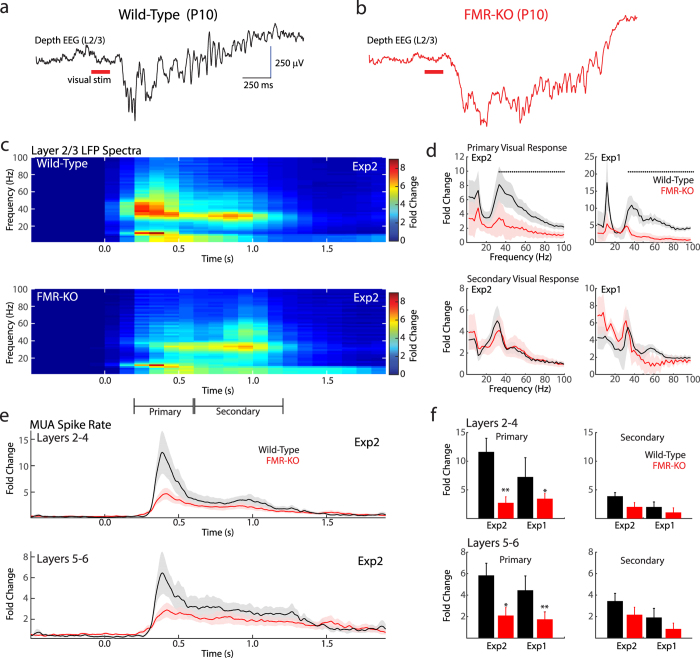
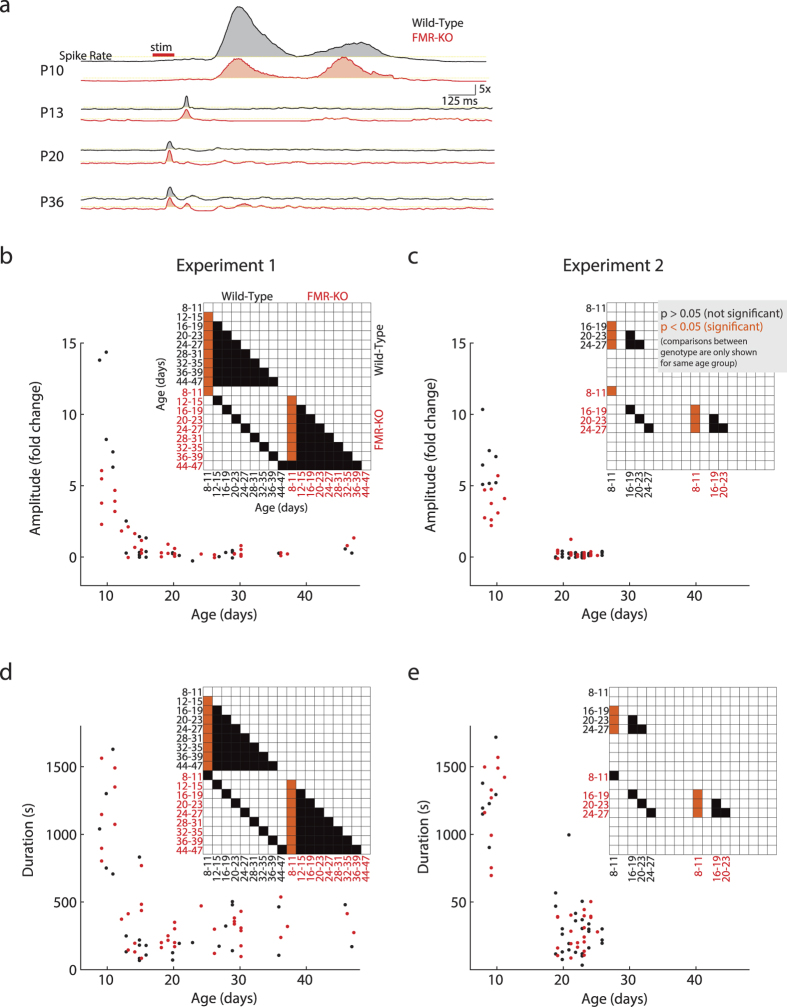
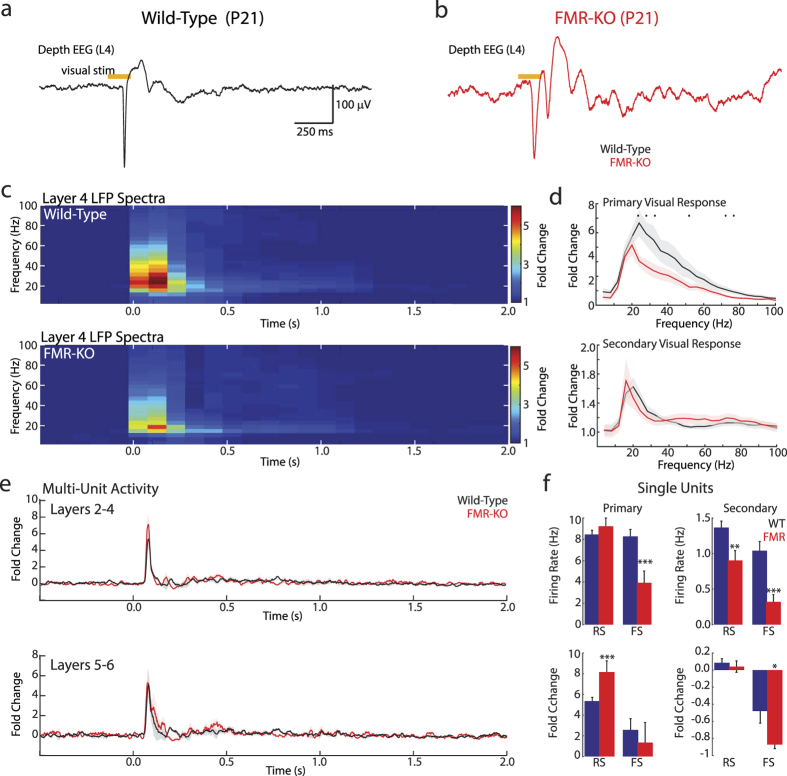
Fragile X syndrome (FXS) is characterized by sensory hyper-sensitivity, and animal models suggest that neuronal hyper-excitability contributes to this phenotype. To understand how sensory dysfunction develops in FXS, we used the rat model (FMR-KO) to quantify the maturation of cortical visual responses from the onset of responsiveness prior to eye-opening, through age equivalents of human juveniles. Rather than hyper-excitability, visual responses before eye-opening had reduced spike rates and an absence of early gamma oscillations, a marker for normal thalamic function at this age. Despite early hypo-excitability, the developmental trajectory of visual responses in FMR-KO rats was normal, and showed the expected loss of visually evoked bursting at the same age as wild-type, two days before eye-opening. At later ages, during the third and fourth post-natal weeks, signs of mild hyper-excitability emerged. These included an increase in the visually-evoked firing of regular spiking, presumptive excitatory, neurons, and a reduced firing of fast-spiking, presumptive inhibitory, neurons. Our results show that early network changes in the FMR-KO rat arise at ages equivalent to fetal humans and have consequences for excitability that are opposite those found in adults. This suggests identification and treatment should begin early, and be tailored in an age-appropriate manner.
Figures
References
-
- Abitbol M. et al. Nucleus basalis magnocellularis and hippocampus are the major sites of FMR-1 expression in the human fetal brain. Nat. Genet. 4, 147–153 (1993). - PubMed
-
- Wijetunge L. S., Chattarji S., Wyllie D. J. & Kind P. C. Fragile X syndrome: from targets to treatments. Neuropharmacology 68, 83–96 (2013). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
