

Lack of p53 Augments Antitumor Functions in Cytolytic T Cells
- PMID: 27466285
- PMCID: PMC5026612
- DOI: 10.1158/0008-5472.CAN-15-1798
Lack of p53 Augments Antitumor Functions in Cytolytic T Cells
Abstract
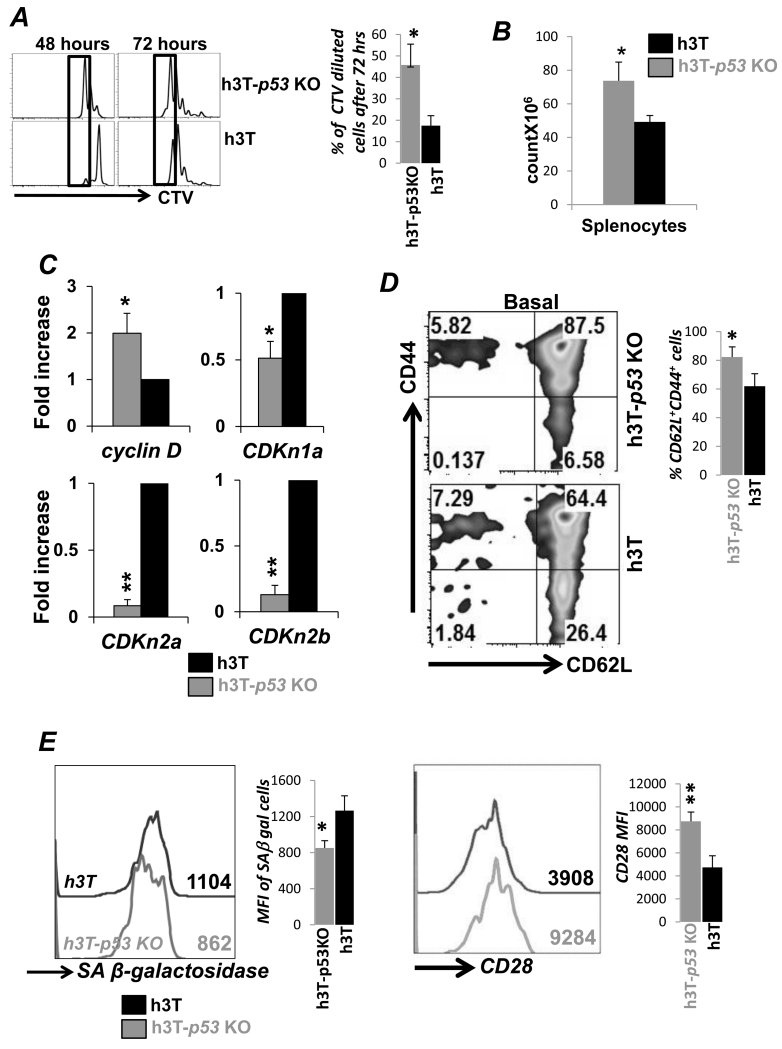
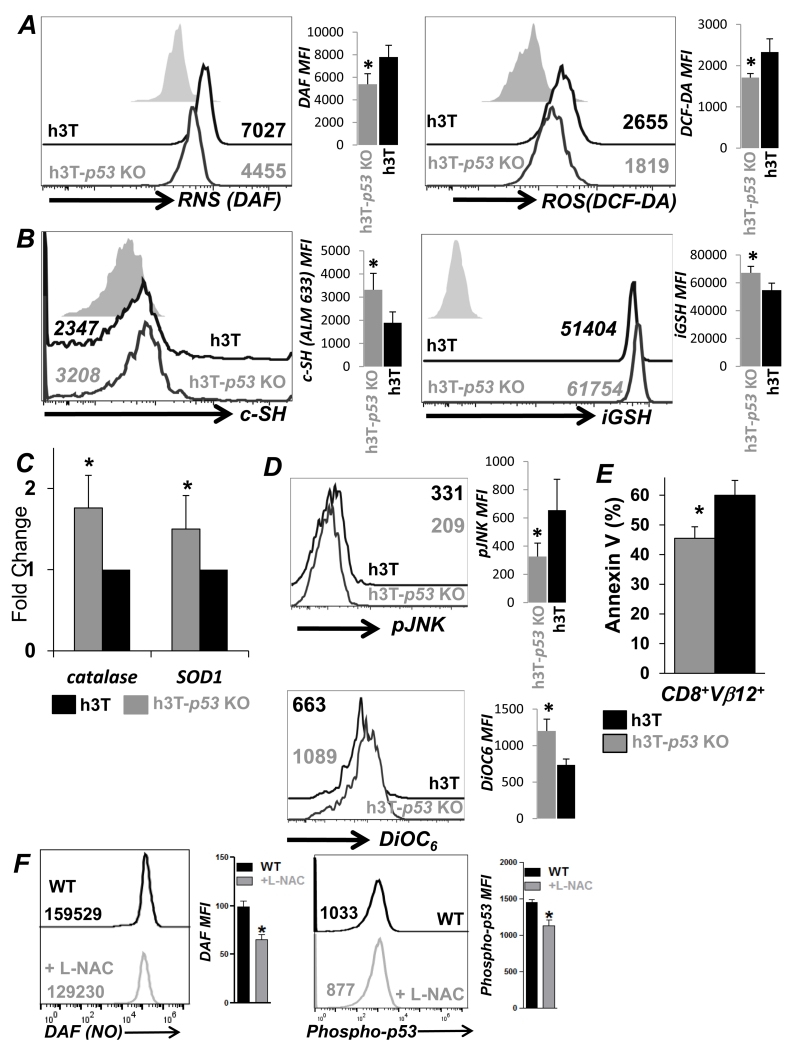
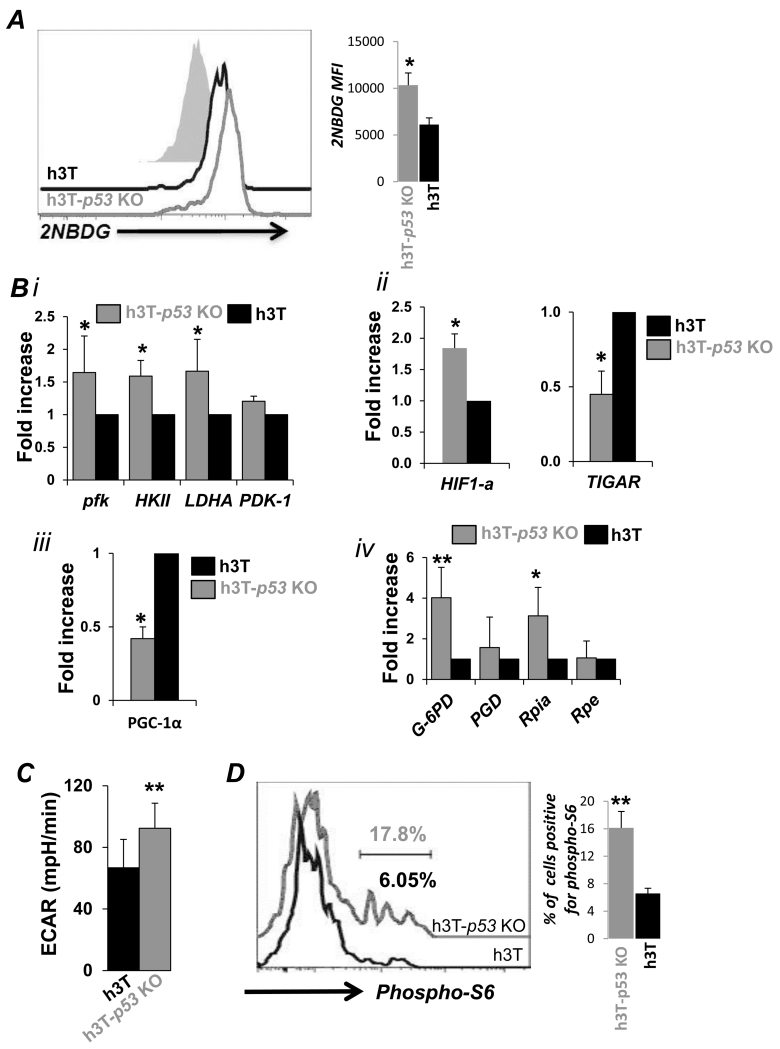
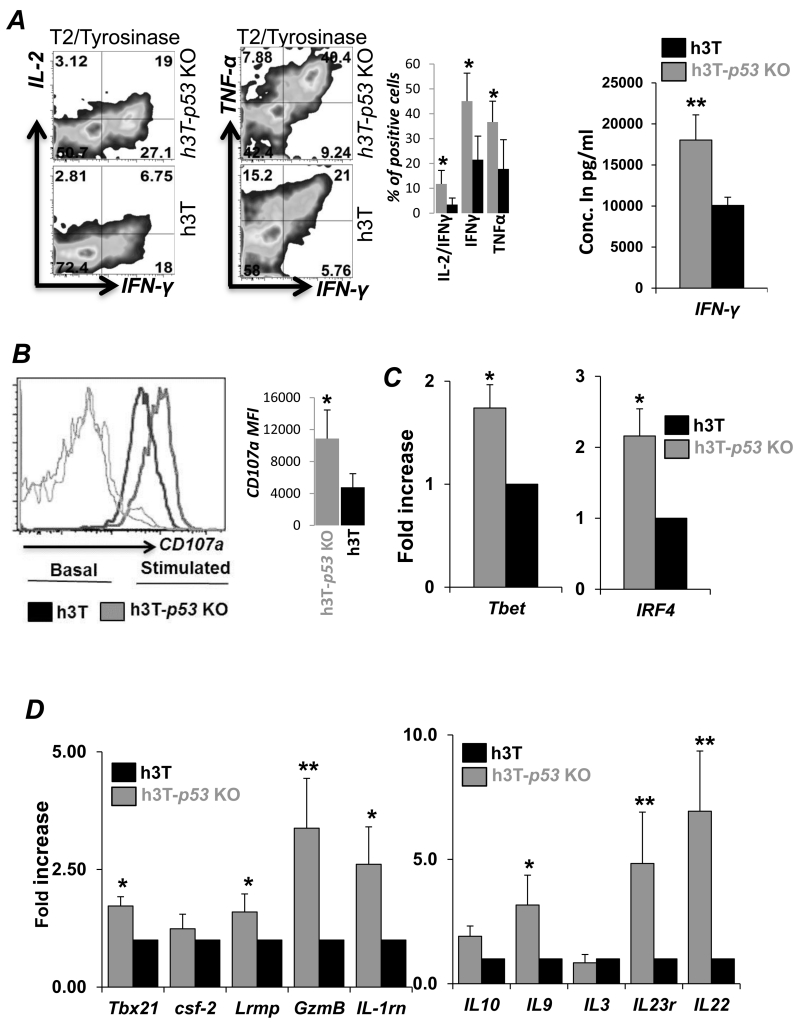
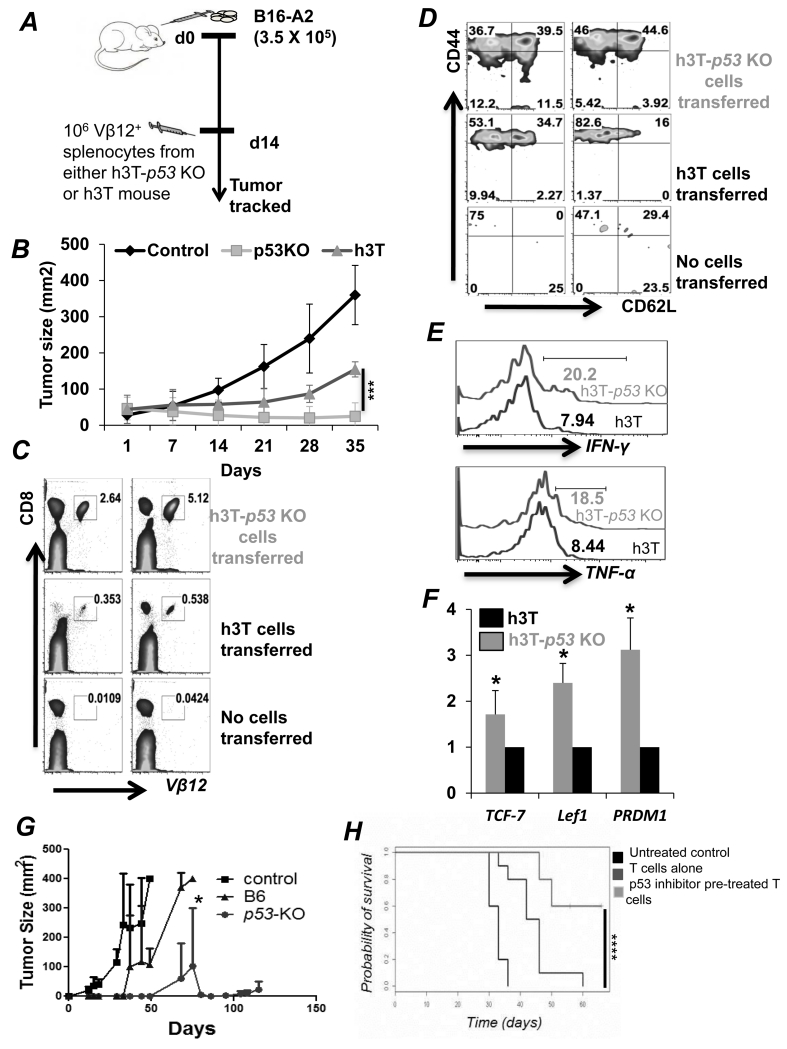
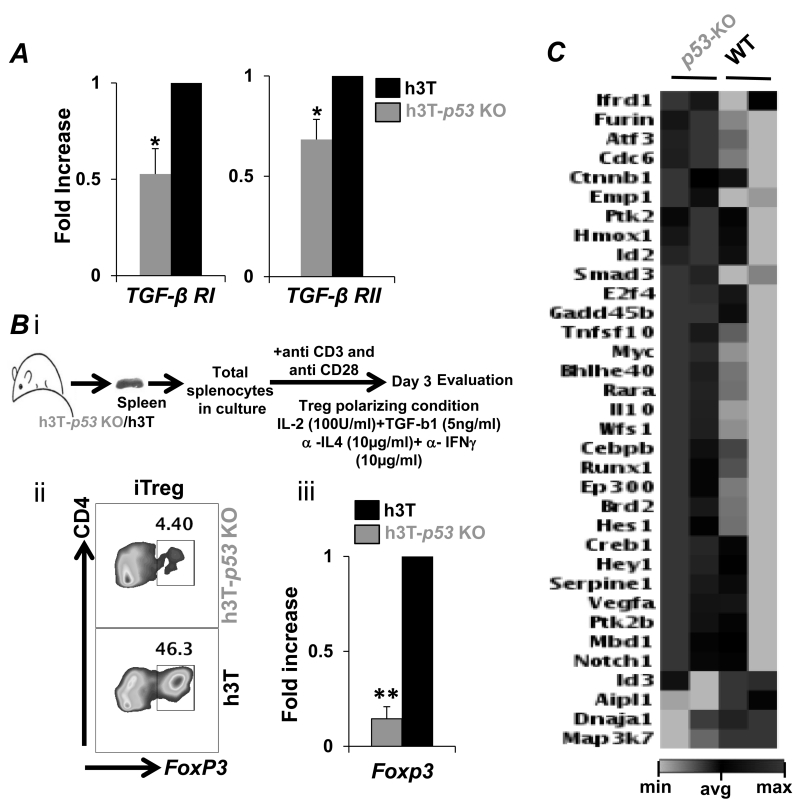
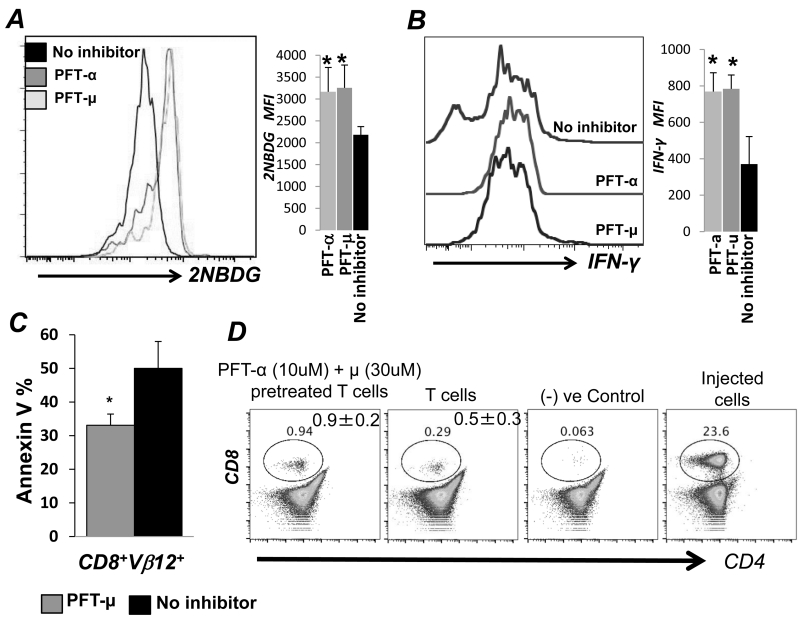
Repetitive stimulation of T-cell receptor (TCR) with cognate antigen results in robust proliferation and expansion of the T cells, and also imprints them with replicative senescence signatures. Our previous studies have shown that life-span and antitumor function of T cells can be enhanced by inhibiting reactive oxygen species (ROS) or intervening with ROS-dependent JNK activation that leads to its activation-induced cell death. Because tumor suppressor protein p53 is also a redox active transcription factor that regulates cellular ROS generation that triggers downstream factor-mediating apoptosis, we determined if p53 levels could influence persistence and function of tumor-reactive T cells. Using h3T TCR transgenic mice, with human tyrosinase epitope-reactive T cells developed on p53 knockout (KO) background, we determined its role in regulating antitumor T-cell function. Our data show that as compared with h3T cells, h3T-p53 KO T cells exhibited enhanced glycolytic commitment that correlated with increased proliferation, IFNγ secretion, cytolytic capacity, expression of stemness gene signature, and decreased TGF-β signaling. This increased effector function correlated to the improved control of subcutaneously established murine melanoma after adoptive transfer of p53-KO T cells. Pharmacological inhibition of human TCR-transduced T cells using a combination of p53 inhibitors also potentiated the T-cell effector function and improved persistence. Thus, our data highlight the key role of p53 in regulating the tumor-reactive T-cell response and that targeting this pathway could have potential translational significance in adoptive T-cell therapy. Cancer Res; 76(18); 5229-40. ©2016 AACR.
©2016 American Association for Cancer Research.
Figures
References
-
- Zuckerman V, Wolyniec K, Sionov RV, Haupt S, Haupt Y. Tumour suppression by p53: the importance of apoptosis and cellular senescence. The Journal of pathology. 2009;219(1):3–15. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
