

Characterization of Simian Immunodeficiency Virus Variants Anatomically Compartmentalized in Plasma and Milk in Chronically Infected African Green Monkeys
- PMID: 27466415
- PMCID: PMC5021398
- DOI: 10.1128/JVI.00701-16
Characterization of Simian Immunodeficiency Virus Variants Anatomically Compartmentalized in Plasma and Milk in Chronically Infected African Green Monkeys
Abstract
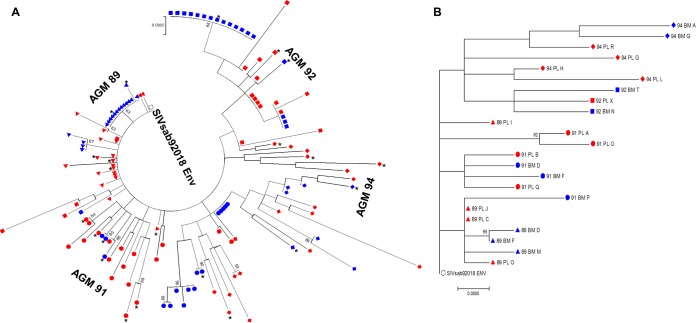
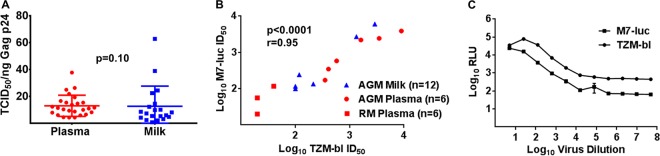
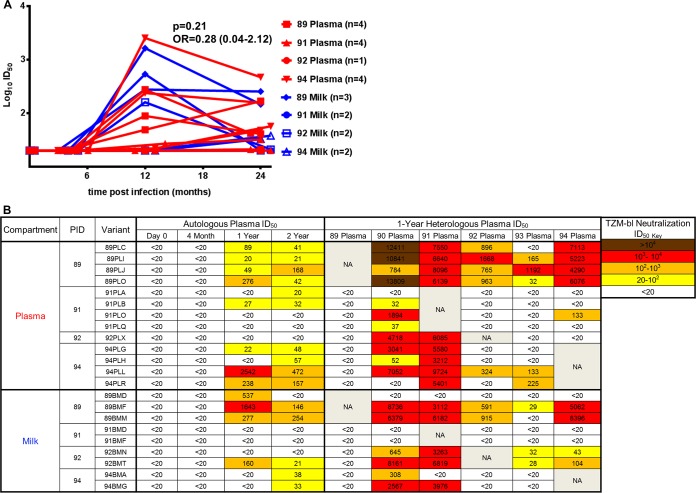
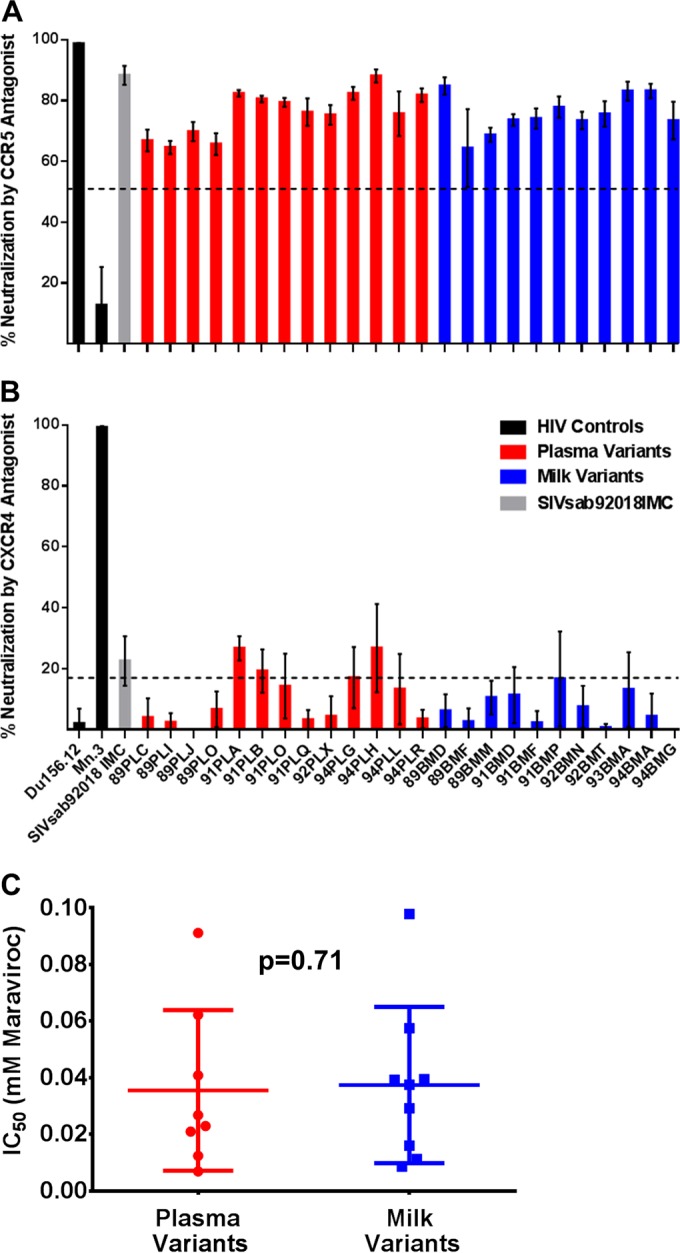
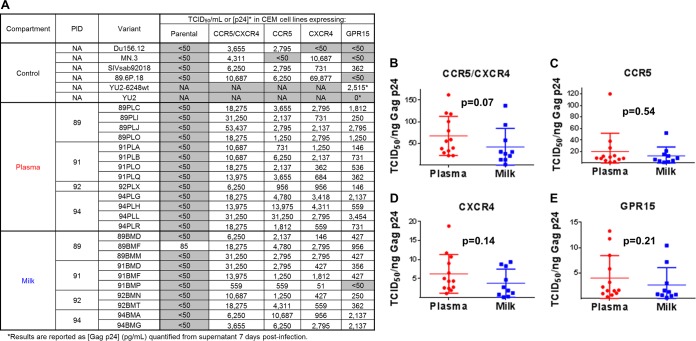
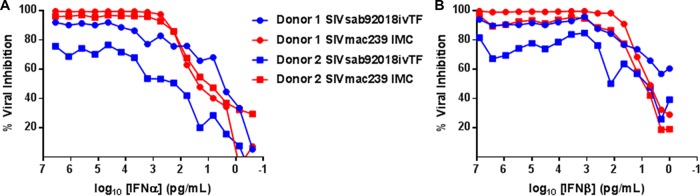
Unlike human immunodeficiency virus type 1 (HIV-1)-infected humans, African-origin, natural simian immunodeficiency virus (SIV) hosts, such as African green monkeys (AGMs), sustain nonpathogenic SIV infections and rarely vertically transmit SIV to their infants. Interestingly, chronically SIV-infected AGMs have anatomically compartmentalized SIV variants in plasma and milk, whereas humans and SIV-infected rhesus monkeys (RMs), Asian-origin nonnatural SIV hosts, do not exhibit this compartmentalization. Thus, it is possible that AGM SIV populations in milk have unique phenotypic features that contribute to the low postnatal transmission rates observed in this natural host species. In this study, we explored this possibility by characterizing the infectivity, tropism, and neutralization susceptibility of plasma and milk SIVsab env variants isolated from chronically SIVsab92018ivTF-infected AGMs. AGM plasma and milk SIVsab env pseudovirus variants exhibited similar infectivities, neutralization susceptibilities to autologous and heterologous plasma, and chemokine coreceptor usages for cell entry, suggesting similar abilities to initiate infection in a new host. We also assessed the cytokine milieu in SIV-infected AGM milk and compared it to that of SIV-infected RMs. MIP-1β, granulocyte colony-stimulating factor (G-CSF), interleukin-12/23 (IL-12/23), and IL-13 trended significantly higher in SIV-infected AGM milk than in that of RMs, while IL-18 and IL-6 trended significantly higher in SIV-infected RM milk than in that of AGMs. Taken together, our findings imply that nonviral maternal factors, such as the cytokine milieu, rather than unique characteristics of SIV populations in the milk contribute to the low postnatal transmission rates observed in AGMs.
Importance: Due to the ongoing global incidence of pediatric HIV-1 infections, including many that occur via breastfeeding, development of effective vaccine strategies capable of preventing vertical HIV transmission through breastfeeding remains an important goal. Unlike HIV-1-infected humans, African green monkeys (AGMs), the natural SIV host species, sustain nonpathogenic SIV infections, rarely transmit the virus postnatally to their infants, and exhibit anatomically compartmentalized SIV populations in milk and plasma. Identifying unique features of the anatomically compartmentalized milk SIV populations could enhance our understanding of how AGMs may have evolved to avoid transmission through breastfeeding. While this study identified limited phenotypic distinctions between AGM plasma and milk SIV populations, potential differences in milk cytokine profiles of natural and nonnatural SIV hosts were observed. These findings imply the potential importance of nonviral factors in natural SIV host species, such as innate SIV/HIV immune factors in milk, as a means of naturally preventing vertical transmission.
Copyright © 2016, American Society for Microbiology. All Rights Reserved.
Figures
Similar articles
-
Rapid Development of gp120-Focused Neutralizing B Cell Responses during Acute Simian Immunodeficiency Virus Infection of African Green Monkeys.J Virol. 2015 Sep;89(18):9485-98. doi: 10.1128/JVI.01564-15. Epub 2015 Jul 8. J Virol. 2015. PMID: 26157116 Free PMC article.
-
High cell-free virus load and robust autologous humoral immune responses in breast milk of simian immunodeficiency virus-infected african green monkeys.J Virol. 2011 Sep;85(18):9517-26. doi: 10.1128/JVI.00796-11. Epub 2011 Jul 6. J Virol. 2011. PMID: 21734053 Free PMC article.
-
Lack of B cell dysfunction is associated with functional, gp120-dominant antibody responses in breast milk of simian immunodeficiency virus-infected African green monkeys.J Virol. 2013 Oct;87(20):11121-34. doi: 10.1128/JVI.01887-13. Epub 2013 Aug 7. J Virol. 2013. PMID: 23926338 Free PMC article.
-
What can natural infection of African monkeys with simian immunodeficiency virus tell us about the pathogenesis of AIDS?AIDS Rev. 2004 Jan-Mar;6(1):40-53. AIDS Rev. 2004. PMID: 15168740 Review.
-
SIVagm: genetic and biological features associated with replication.Front Biosci. 2003 Sep 1;8:d1170-85. doi: 10.2741/1130. Front Biosci. 2003. PMID: 12957815 Review.
References
-
- UNAIDS. 2014. 2014 progress report on the global plan towards the elimination of new HIV infections among children by 2015 and keeping their mothers alive. UNAIDS, Washington, DC.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
