

High Dietary Selenium Intake Alters Lipid Metabolism and Protein Synthesis in Liver and Muscle of Pigs
- PMID: 27466604
- PMCID: PMC4997278
- DOI: 10.3945/jn.116.229955
High Dietary Selenium Intake Alters Lipid Metabolism and Protein Synthesis in Liver and Muscle of Pigs
Abstract
Background: Prolonged high intakes of dietary selenium have been shown to induce gestational diabetes in rats and hyperinsulinemia in pigs.
Objective: Two experiments were conducted to explore metabolic and molecular mechanisms for the diabetogenic potential of high dietary selenium intakes in pigs.
Methods: In Expt. 1, 16 Yorkshire-Landrace-Hampshire crossbred pigs (3 wk old, body weight = 7.5 ± 0.81 kg, 50% males and 50% females) were fed a corn-soybean meal basal diet supplemented with 0.3 or 1.0 mg Se/kg (as selenium-enriched yeast for 6 wk). In Expt. 2, 12 pigs of the same crossbreed (6 wk old, body weight = 16.0 ± 1.8 kg) were fed a similar basal diet supplemented with 0.3 or 3.0 mg Se/kg for 11 wk. Biochemical and gene and protein expression profiles of lipid and protein metabolism and selenoproteins in plasma, liver, muscle, and adipose tissues were analyzed.
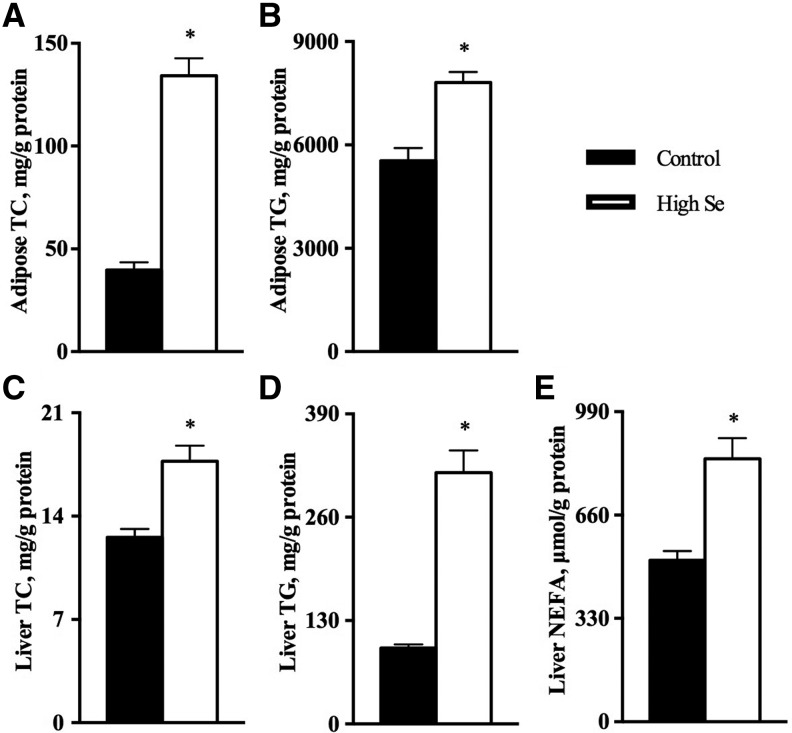
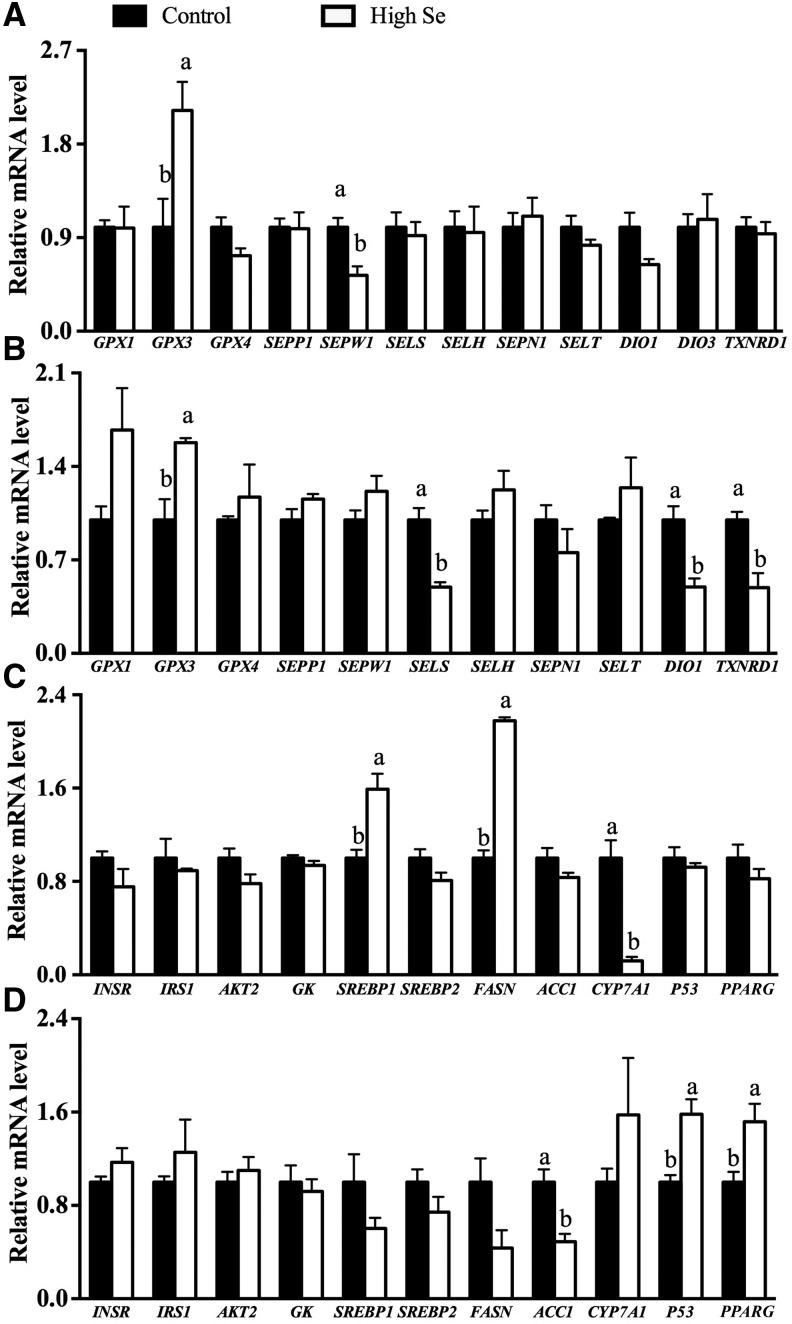
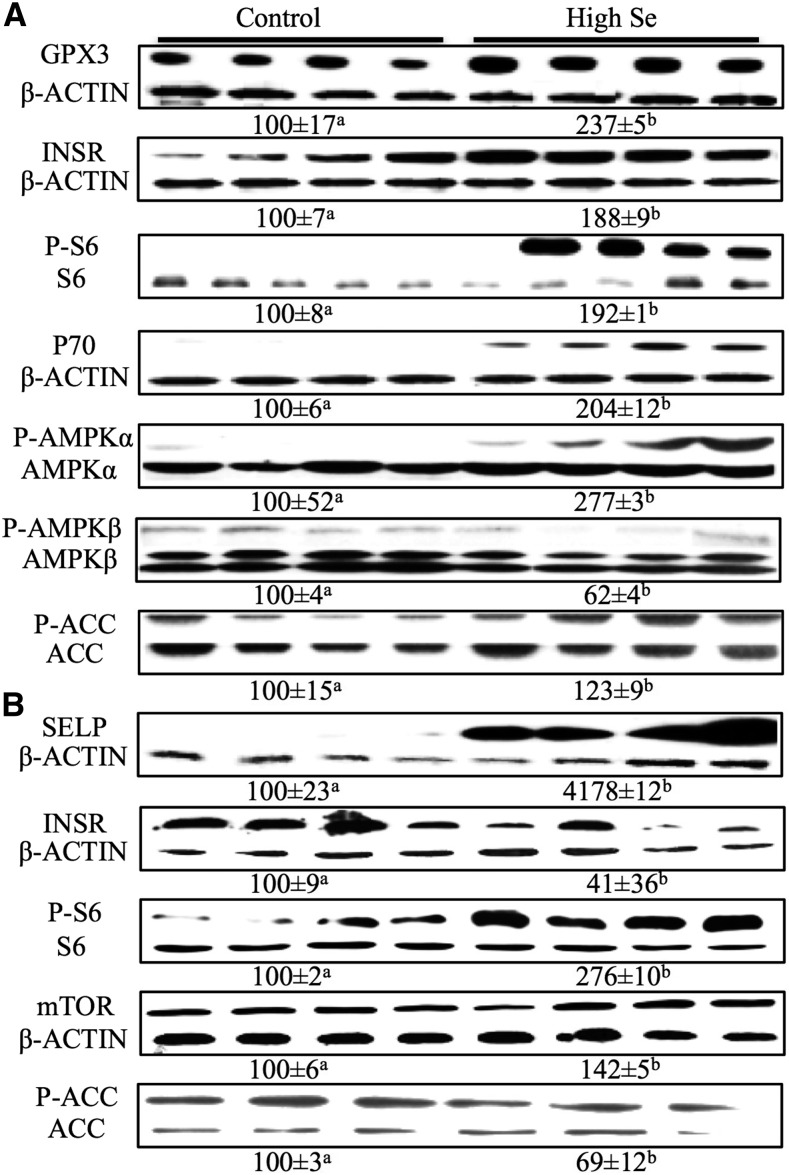
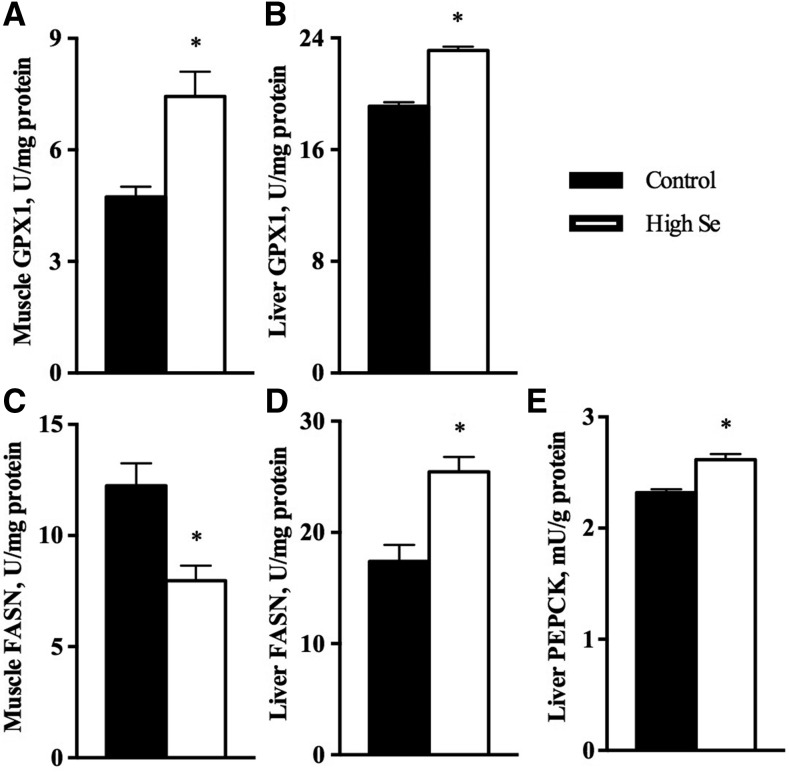
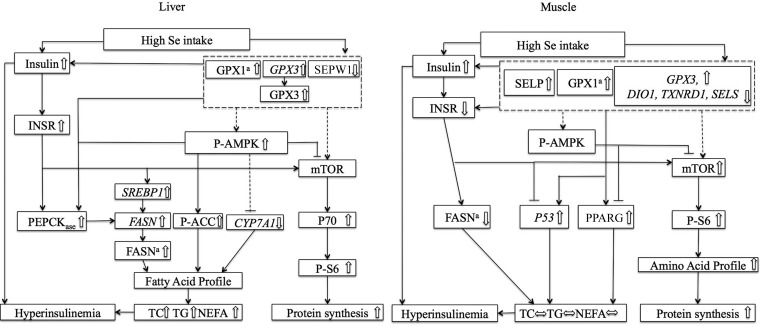
Results: In Expt. 1, the 1-mg-Se/kg diet did not affect body weight or plasma concentrations of glucose and nonesterified fatty acids. In Expt. 2, the 3-mg-Se/kg diet, compared with the 0.3-mg-Se/kg diet, increased (P < 0.05) concentrations of plasma insulin (0.2 compared with 0.4 ng/mL), liver and adipose lipids (41% to 2.4-fold), and liver and muscle protein (10-14%). In liver, the 3-mg-Se/kg diet upregulated (P < 0.05) the expression, activity, or both of key factors related to gluconeogenesis [phosphoenolpyruvate carboxykinase (PEPCK); 13%], lipogenesis [sterol regulatory element binding protein 1 (SREBP1), acetyl-coenzyme A carboxylase (ACC), and fatty acid synthase (FASN); 46-90%], protein synthesis [insulin receptor (INSR), P70 ribosomal protein S6 kinase (P70), and phosphorylated ribosomal protein S6 (P-S6); 88-105%], energy metabolism [AMP-activated protein kinase (AMPK); up to 2.8-fold], and selenoprotein glutathione peroxidase 3 (GPX3; 1.4-fold) and suppressed (P < 0.05) mRNA levels of lipolysis gene cytochrome P450, family 7, subfamily A, polypeptide 1 (CYP7A1; 88%) and selenoprotein gene selenoprotein W1 (SEPW1; 46%). In muscle, the 3-mg-Se/kg diet exerted no effect on the lipid profiles but enhanced (P < 0.05) expression of P-S6 and mammalian target of rapamycin (mTOR; 42-176%; protein synthesis); selenoprotein P (SELP; 40-fold); and tumor suppressor protein 53 (P53) and peroxisome proliferator-activated receptor γ (PPARG; 52-58%; lipogenesis) and suppressed (P < 0.05) expression of INSR (59%; insulin signaling); selenoprotein S (SELS); deiodinases, iodothyronine, type I (DIO1); and thioredoxin reductase 1 (TXNRD1; 50%; selenoproteins); and ACC1 and FASN (35-51%; lipogenesis).
Conclusion: Our research showed novel roles, to our best knowledge, and mechanisms of high selenium intakes in regulating the metabolism of protein, along with that of lipid, in a tissue-specific fashion in pigs.
Keywords: AMPK; Se; lipid; protein; selenoprotein.
© 2016 American Society for Nutrition.
Conflict of interest statement
2 Author disclosures: Z Zhao, M Barcus, J Kim, KL Lum, C Mills, and XG Lei, no conflicts of interest.
Figures
References
-
- Wang XD, Vatamaniuk MZ, Wang SK, Roneker CA, Simmons RA, Lei XG. Molecular mechanisms for hyperinsulinaemia induced by overproduction of selenium-dependent glutathione peroxidase-1 in mice. Diabetologia 2008;51:1515–24. - PubMed
-
- Pinto A, Juniper DT, Sanil M, Morgan L, Clark L, Sies H, Rayman MP, Steinbrenner H. Supranutritional selenium induces alterations in molecular targets related to energy metabolism in skeletal muscle and visceral adipose tissue of pigs. J Inorg Biochem 2012;114:47–54. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
