

Selection on a Subunit of the NURF Chromatin Remodeler Modifies Life History Traits in a Domesticated Strain of Caenorhabditis elegans
- PMID: 27467070
- PMCID: PMC4965130
- DOI: 10.1371/journal.pgen.1006219
Selection on a Subunit of the NURF Chromatin Remodeler Modifies Life History Traits in a Domesticated Strain of Caenorhabditis elegans
Abstract
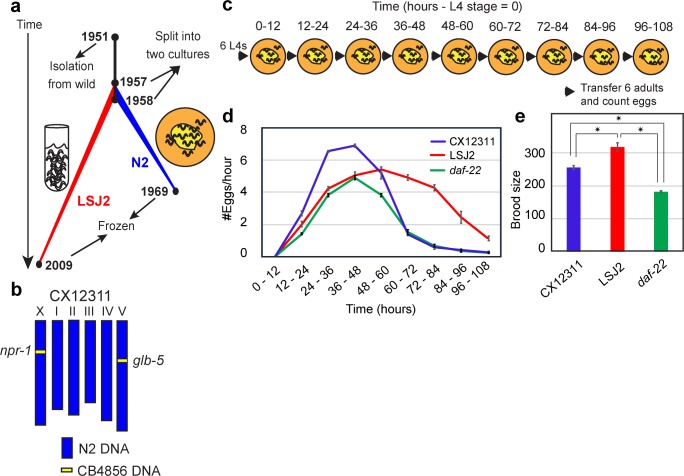
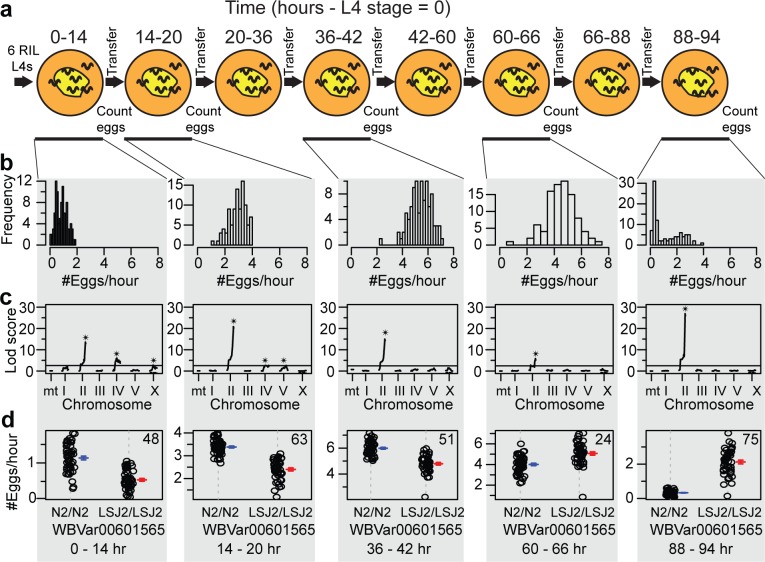
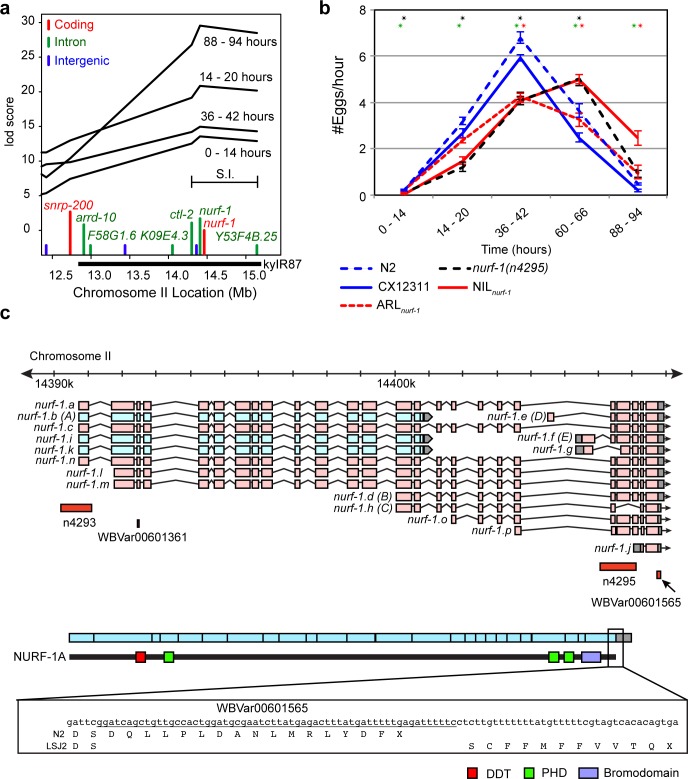
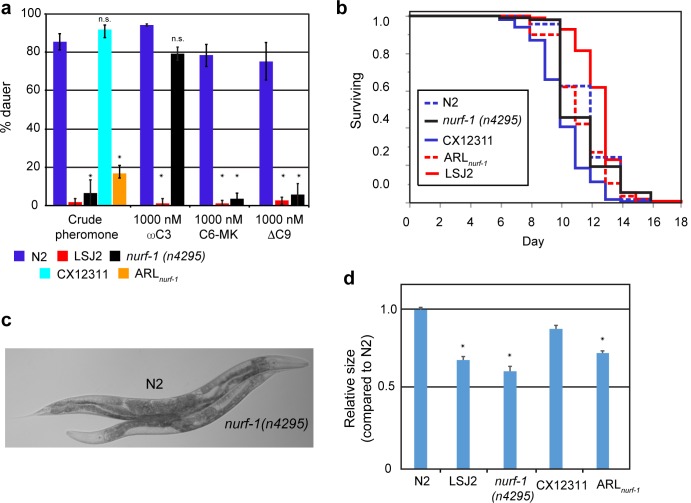
Evolutionary life history theory seeks to explain how reproductive and survival traits are shaped by selection through allocations of an individual's resources to competing life functions. Although life-history traits evolve rapidly, little is known about the genetic and cellular mechanisms that control and couple these tradeoffs. Here, we find that two laboratory-adapted strains of C. elegans descended from a single common ancestor that lived in the 1950s have differences in a number of life-history traits, including reproductive timing, lifespan, dauer formation, growth rate, and offspring number. We identified a quantitative trait locus (QTL) of large effect that controls 24%-75% of the total trait variance in reproductive timing at various timepoints. Using CRISPR/Cas9-induced genome editing, we show this QTL is due in part to a 60 bp deletion in the 3' end of the nurf-1 gene, which is orthologous to the human gene encoding the BPTF component of the NURF chromatin remodeling complex. Besides reproduction, nurf-1 also regulates growth rate, lifespan, and dauer formation. The fitness consequences of this deletion are environment specific-it increases fitness in the growth conditions where it was fixed but decreases fitness in alternative laboratory growth conditions. We propose that chromatin remodeling, acting through nurf-1, is a pleiotropic regulator of life history trade-offs underlying the evolution of multiple traits across different species.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Life history in Caenorhabditis elegans: from molecular genetics to evolutionary ecology.Genetics. 2024 Nov 6;228(3):iyae151. doi: 10.1093/genetics/iyae151. Genetics. 2024. PMID: 39422376 Free PMC article. Review.
-
Evolution of Yin and Yang isoforms of a chromatin remodeling subunit precedes the creation of two genes.Elife. 2019 Sep 9;8:e48119. doi: 10.7554/eLife.48119. Elife. 2019. PMID: 31498079 Free PMC article.
-
Natural Variation and Genetic Determinants of Caenorhabditis elegans Sperm Size.Genetics. 2019 Oct;213(2):615-632. doi: 10.1534/genetics.119.302462. Epub 2019 Aug 8. Genetics. 2019. PMID: 31395653 Free PMC article.
-
Modeling of a negative feedback mechanism explains antagonistic pleiotropy in reproduction in domesticated Caenorhabditis elegans strains.PLoS Genet. 2017 May 11;13(5):e1006769. doi: 10.1371/journal.pgen.1006769. eCollection 2017 May. PLoS Genet. 2017. PMID: 28493873 Free PMC article.
-
[The SWI/SNF chromatin-remodeling complex regulates asymmetric cell division in C. elegans].Tanpakushitsu Kakusan Koso. 2005 May;50(6 Suppl):569-74. Tanpakushitsu Kakusan Koso. 2005. PMID: 15926482 Review. Japanese. No abstract available.
Cited by
-
Chromatin: the old and young of it.Front Mol Biosci. 2023 Oct 9;10:1270285. doi: 10.3389/fmolb.2023.1270285. eCollection 2023. Front Mol Biosci. 2023. PMID: 37877123 Free PMC article. Review.
-
Life history in Caenorhabditis elegans: from molecular genetics to evolutionary ecology.Genetics. 2024 Nov 6;228(3):iyae151. doi: 10.1093/genetics/iyae151. Genetics. 2024. PMID: 39422376 Free PMC article. Review.
-
A multi-parent recombinant inbred line population of C. elegans allows identification of novel QTLs for complex life history traits.BMC Biol. 2019 Mar 12;17(1):24. doi: 10.1186/s12915-019-0642-8. BMC Biol. 2019. PMID: 30866929 Free PMC article.
-
The Ancestral Caenorhabditis elegans Cuticle Suppresses rol-1.G3 (Bethesda). 2020 Jul 7;10(7):2385-2395. doi: 10.1534/g3.120.401336. G3 (Bethesda). 2020. PMID: 32423919 Free PMC article.
-
From QTL to gene: C. elegans facilitates discoveries of the genetic mechanisms underlying natural variation.Trends Genet. 2021 Oct;37(10):933-947. doi: 10.1016/j.tig.2021.06.005. Epub 2021 Jul 3. Trends Genet. 2021. PMID: 34229867 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
