

Vaccine-generated lung tissue-resident memory T cells provide heterosubtypic protection to influenza infection
- PMID: 27468427
- PMCID: PMC4959801
- DOI: 10.1172/jci.insight.85832
Vaccine-generated lung tissue-resident memory T cells provide heterosubtypic protection to influenza infection
Abstract
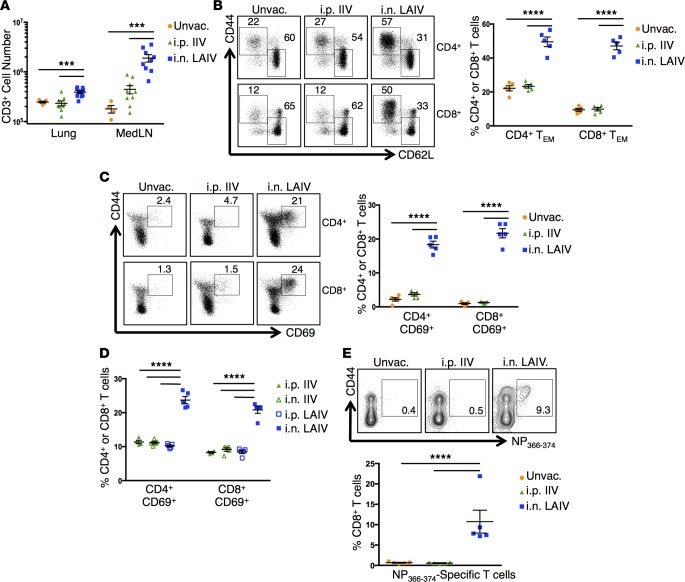
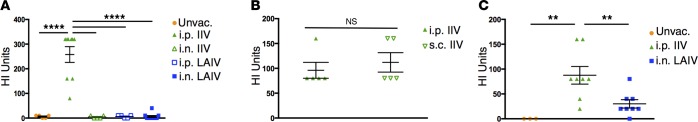
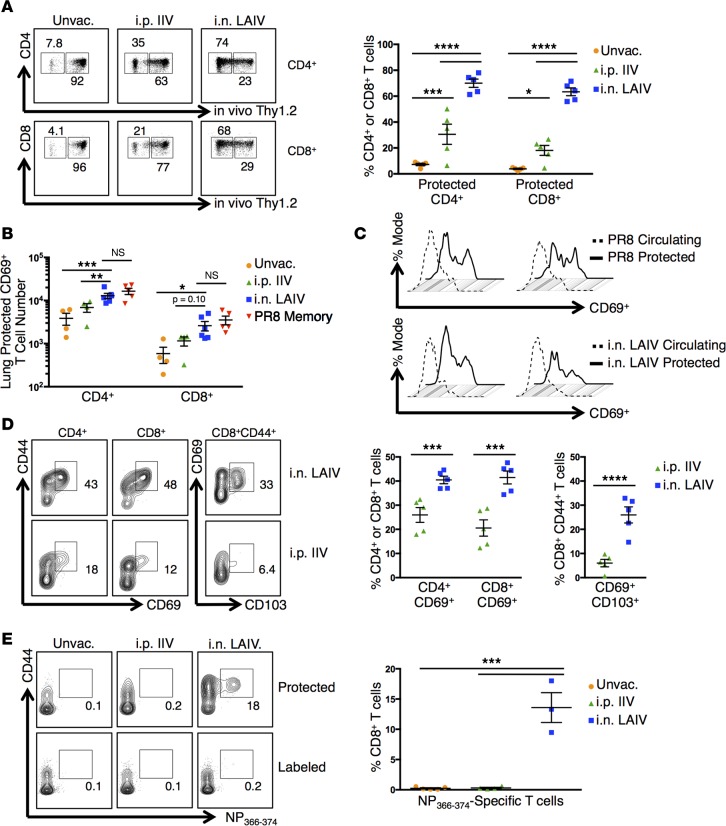
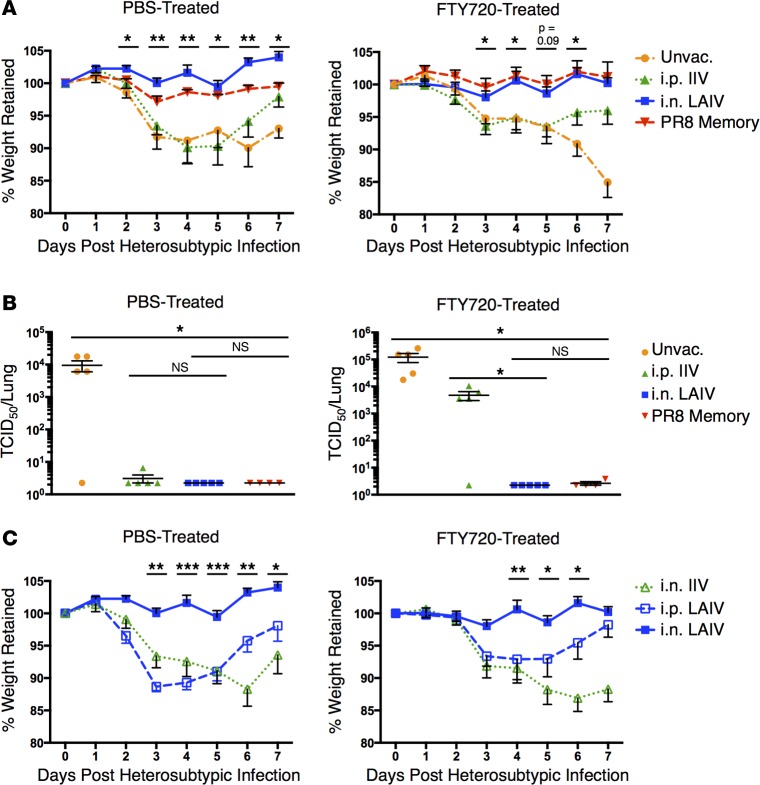
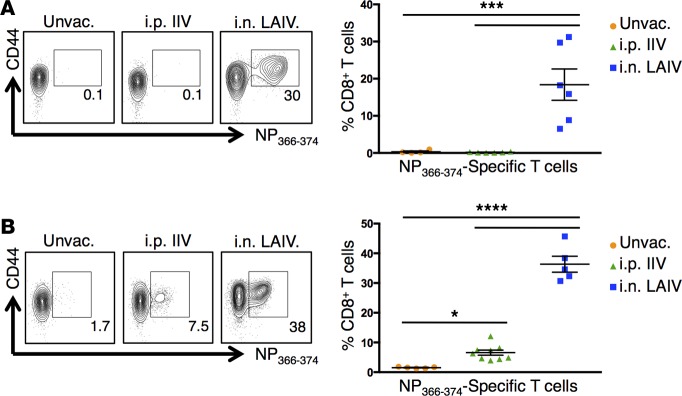
Tissue-resident memory T cells (TRM) are a recently defined, noncirculating subset with the potential for rapid in situ protective responses, although their generation and role in vaccine-mediated immune responses is unclear. Here, we assessed TRM generation and lung-localized protection following administration of currently licensed influenza vaccines, including injectable inactivated influenza virus (IIV, Fluzone) and i.n. administered live-attenuated influenza virus (LAIV, FluMist) vaccines. We found that, while IIV preferentially induced strain-specific neutralizing antibodies, LAIV generated lung-localized, virus-specific T cell responses. Moreover, LAIV but not IIV generated lung CD4+ TRM and virus-specific CD8+ TRM, similar in phenotype to those generated by influenza virus infection. Importantly, these vaccine-generated TRM mediated cross-strain protection, independent of circulating T cells and neutralizing antibodies, which persisted long-term after vaccination. Interestingly, intranasal administration of IIV or injection of LAIV failed to elicit T cell responses or provide protection against viral infection, demonstrating dual requirements for respiratory targeting and a live-attenuated strain to establish TRM. The ability of LAIV to generate lung TRM capable of providing long-term protection against nonvaccine viral strains, as demonstrated here, has important implications for protecting the population against emergent influenza pandemics by direct fortification of lung-specific immunity.
Figures
References
-
- Fiore AE, et al. Prevention and control of influenza with vaccines: recommendations of the Advisory Committee on Immunization Practices (ACIP), 2010. MMWR Recomm Rep. 2010;59(RR-8):1–62. - PubMed
-
- McIntyre AF, Gonzalez-Feliciano AG, Bryan LN, Santibanez TA, Williams WW, Singleton JA. Seasonal influenza vaccination coverage - United States, 2009-10 and 2010-11. MMWR Surveill Summ. 2013;62 Suppl 3(3):65–68. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
