

Atomic-resolution structure of a disease-relevant Aβ(1-42) amyloid fibril
- PMID: 27469165
- PMCID: PMC5003276
- DOI: 10.1073/pnas.1600749113
Atomic-resolution structure of a disease-relevant Aβ(1-42) amyloid fibril
Abstract
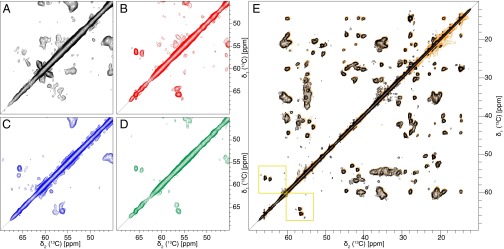
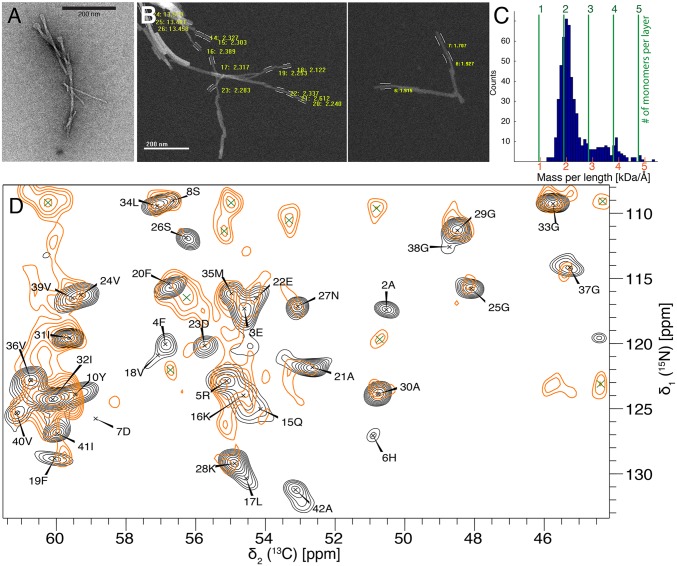
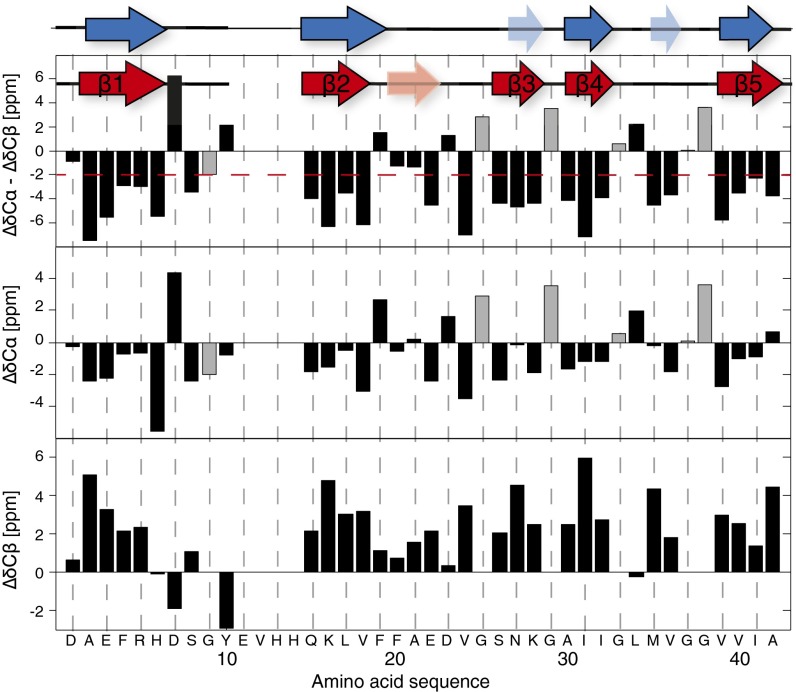
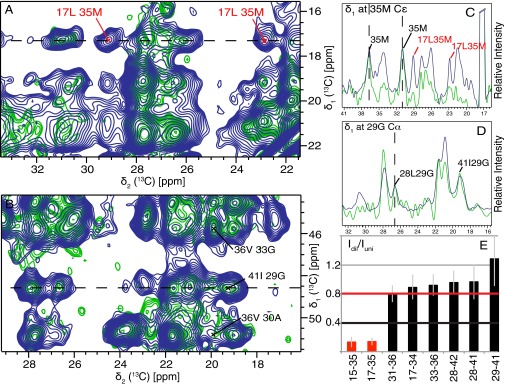
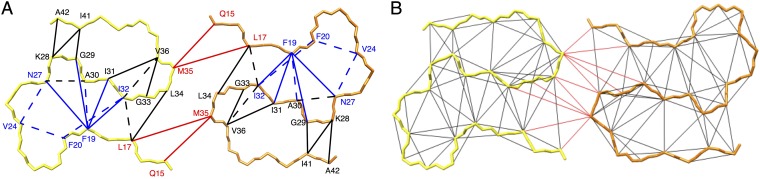
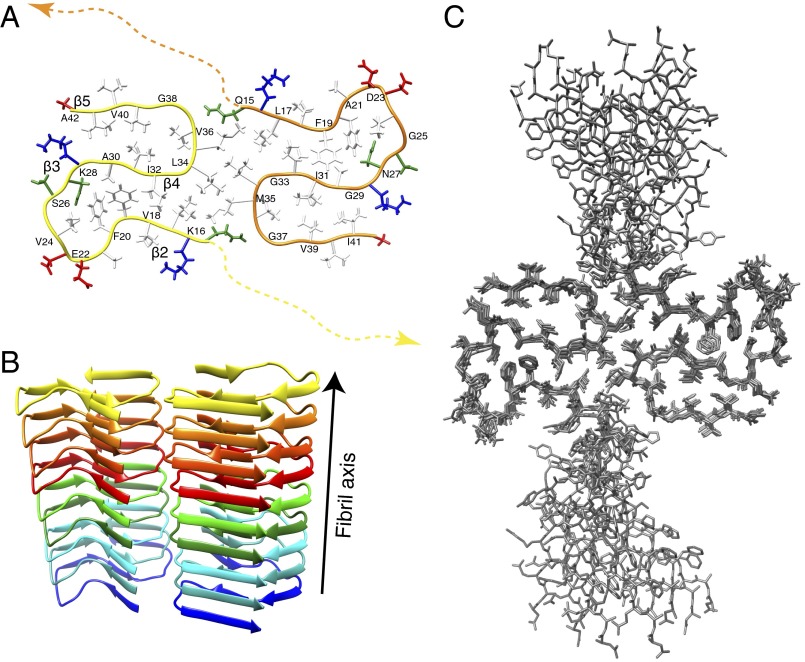
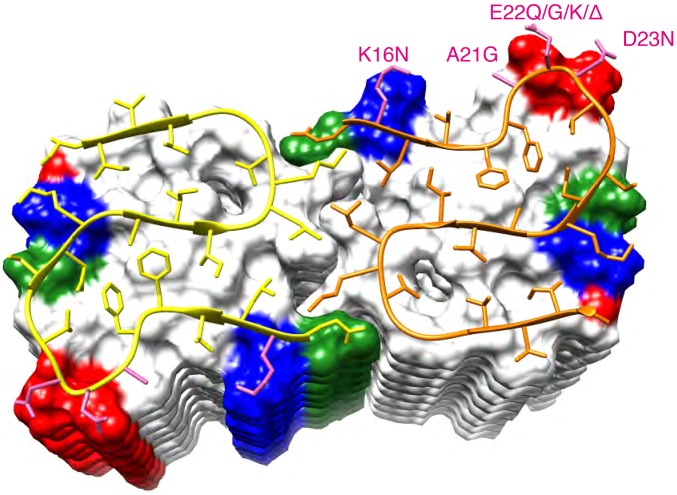
Amyloid-β (Aβ) is present in humans as a 39- to 42-amino acid residue metabolic product of the amyloid precursor protein. Although the two predominant forms, Aβ(1-40) and Aβ(1-42), differ in only two residues, they display different biophysical, biological, and clinical behavior. Aβ(1-42) is the more neurotoxic species, aggregates much faster, and dominates in senile plaque of Alzheimer's disease (AD) patients. Although small Aβ oligomers are believed to be the neurotoxic species, Aβ amyloid fibrils are, because of their presence in plaques, a pathological hallmark of AD and appear to play an important role in disease progression through cell-to-cell transmissibility. Here, we solved the 3D structure of a disease-relevant Aβ(1-42) fibril polymorph, combining data from solid-state NMR spectroscopy and mass-per-length measurements from EM. The 3D structure is composed of two molecules per fibril layer, with residues 15-42 forming a double-horseshoe-like cross-β-sheet entity with maximally buried hydrophobic side chains. Residues 1-14 are partially ordered and in a β-strand conformation, but do not display unambiguous distance restraints to the remainder of the core structure.
Keywords: Alzheimer’s disease; amyloid; protein structure; solid-state NMR.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Implications for Alzheimer's disease of an atomic resolution structure of amyloid-β(1-42) fibrils.Proc Natl Acad Sci U S A. 2016 Aug 23;113(34):9398-400. doi: 10.1073/pnas.1610806113. Epub 2016 Aug 9. Proc Natl Acad Sci U S A. 2016. PMID: 27506787 Free PMC article. No abstract available.
-
Alzheimer's disease: Structure of aggregates revealed.Nature. 2016 Sep 22;537(7621):492-493. doi: 10.1038/nature19470. Epub 2016 Sep 14. Nature. 2016. PMID: 27626376 Free PMC article.
References
-
- Glenner GG, Wong CW. Alzheimer’s disease and Down’s syndrome: Sharing of a unique cerebrovascular amyloid fibril protein. Biochem Biophys Res Commun. 1984;122(3):1131–1135. - PubMed
-
- Kang J, et al. The precursor of Alzheimer’s disease amyloid A4 protein resembles a cell-surface receptor. Nature. 1987;325(6106):733–736. - PubMed
-
- Costanzo M, Zurzolo C. The cell biology of prion-like spread of protein aggregates: Mechanisms and implication in neurodegeneration. Biochem J. 2013;452(1):1–17. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
