

Stanniocalcin-1 attenuates ischemic cardiac injury and response of differentiating monocytes/macrophages to inflammatory stimuli
- PMID: 27469269
- PMCID: PMC5099094
- DOI: 10.1016/j.trsl.2016.06.011
Stanniocalcin-1 attenuates ischemic cardiac injury and response of differentiating monocytes/macrophages to inflammatory stimuli
Abstract
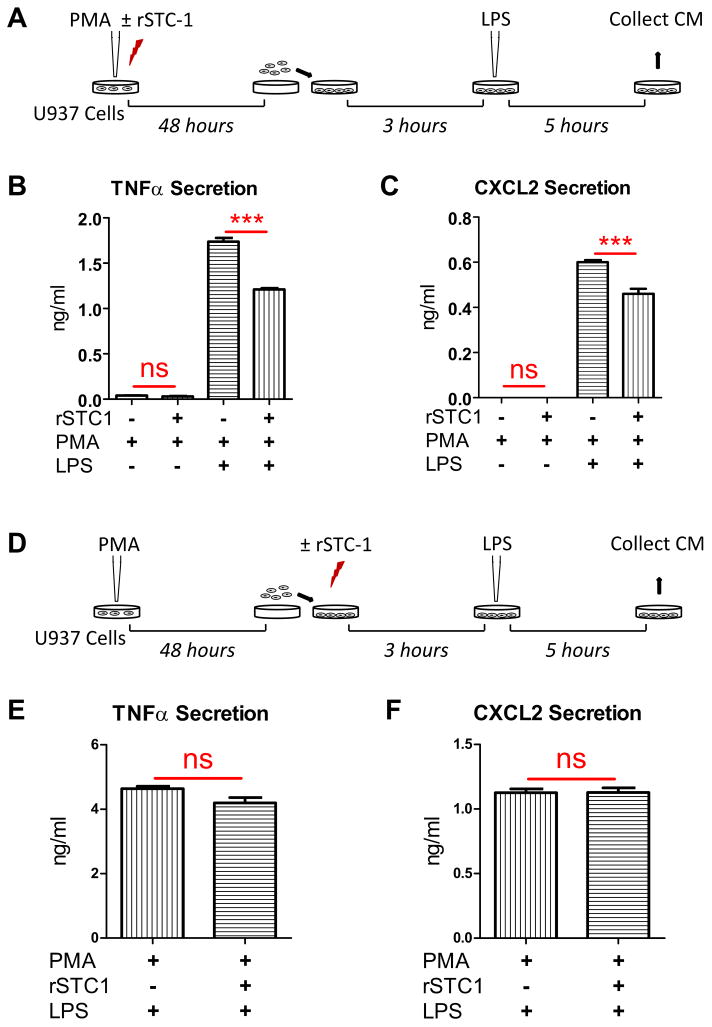
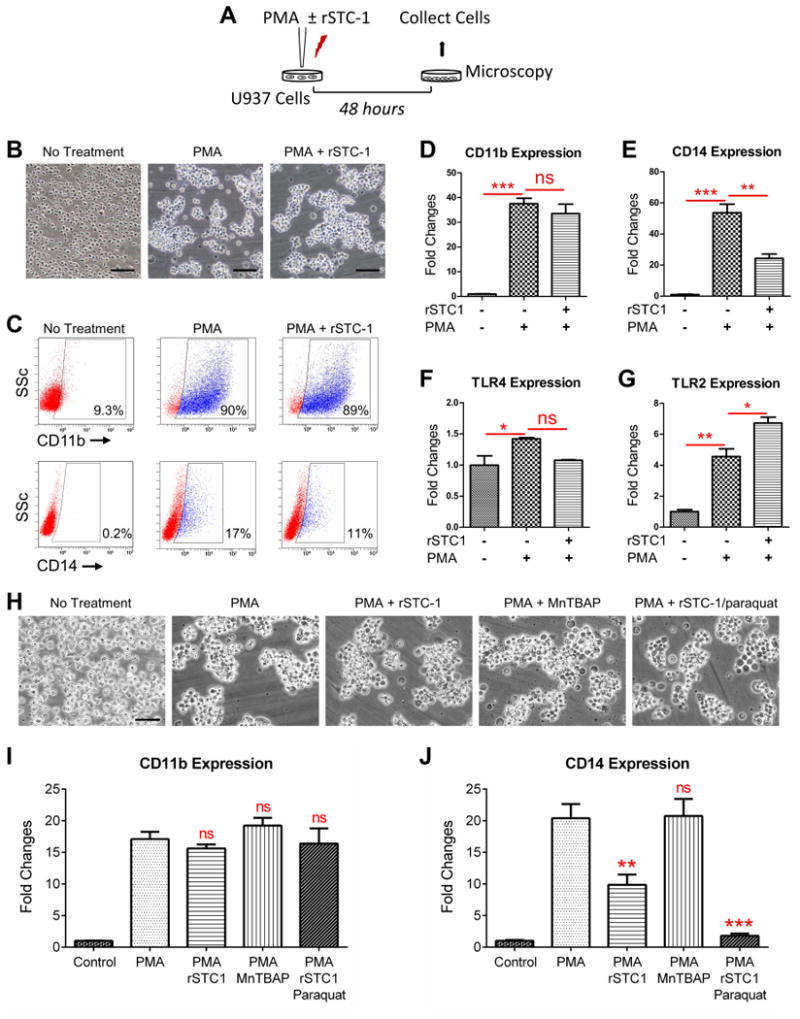
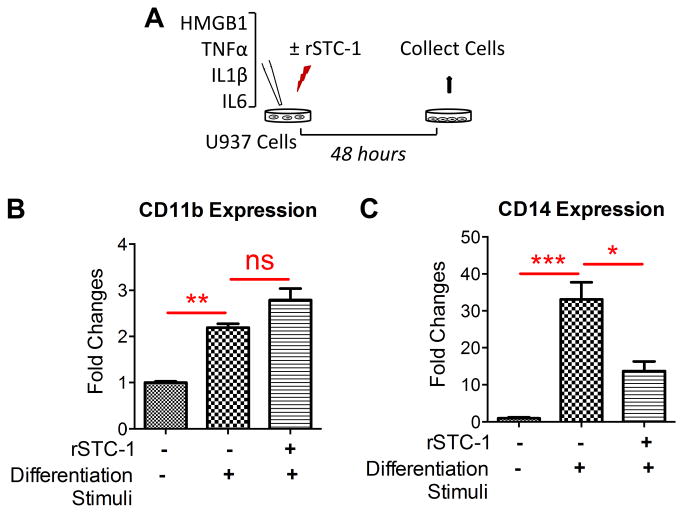
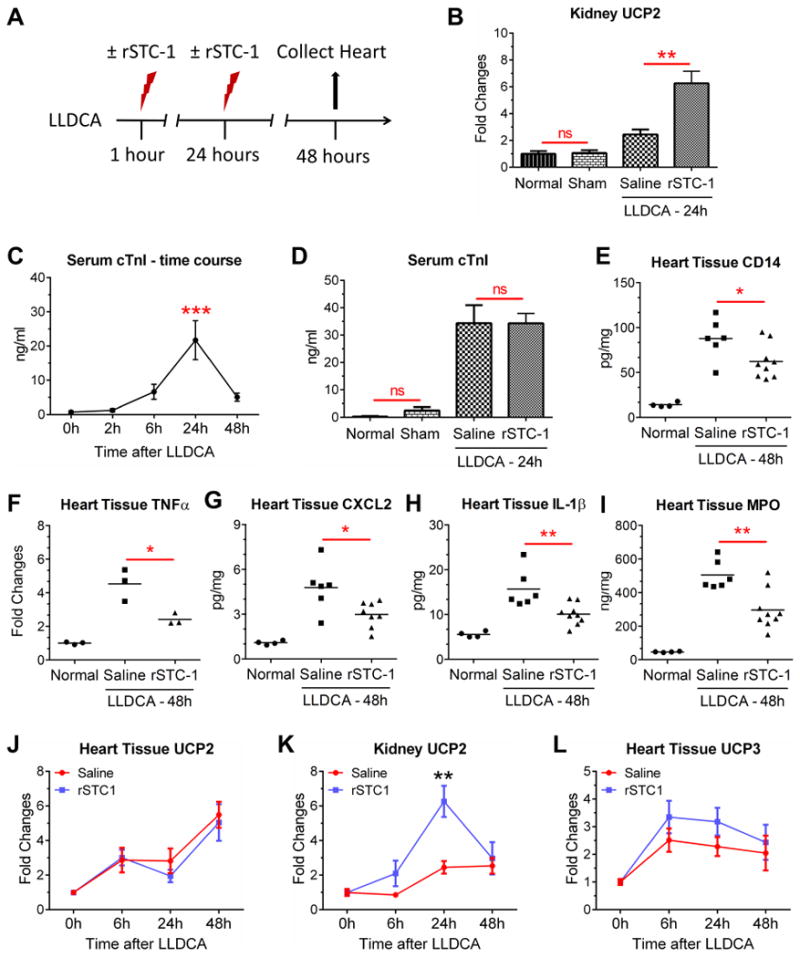
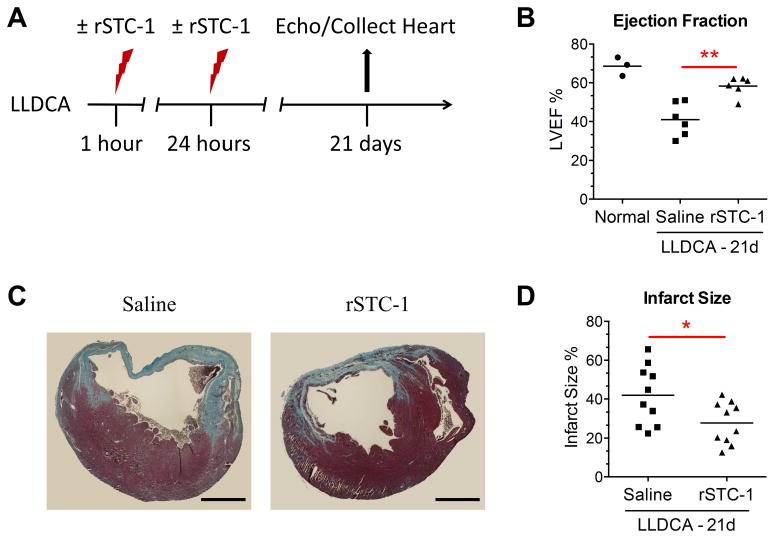
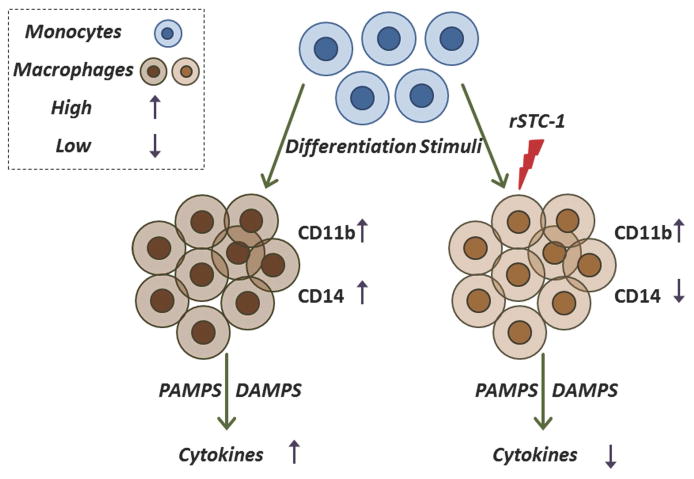
Stanniocalcin-1 (STC-1) is a multifunctional glycoprotein with antioxidant and anti-inflammatory properties. Ischemic myocardial necrosis generates "danger" signals that perpetuate detrimental inflammatory reactions often involving monocyte recruitment and their subsequent differentiation into proinflammatory macrophages. Therefore, we evaluated the effects of recombinant STC-1 (rSTC-1) on monocyte phenotype and in a mouse model of myocardial infarction. Using an established protocol to differentiate human monocytes into macrophages, we demonstrated that rSTC-1 did not alter morphology of the differentiated cells, toll-like receptor (TLR) 4 expression, or expression of the myeloid cell marker CD11b. However, rSTC-1 treatment before differentiation attenuated the rise in the expression of CD14, a TLR4 coreceptor and pathogen sensor that propagates innate immune responses, and suppressed levels of inflammatory cytokines produced by the differentiated cells in response to the CD14-TLR4 ligand lipopolysaccharide. Moreover, rSTC-1 treatment reduced CD14 expression in monocytes stimulated with endogenous danger signals. Interestingly, the effects of rSTC-1 on CD14 expression were not reproduced by a superoxide dismutase mimetic. In mice with induced myocardial infarcts, intravenous administration of rSTC-1 decreased CD14 expression in the heart as well as levels of tumor necrosis factor alpha, C-X-C motif ligand 2, interleukin 1 beta, and myeloperoxidase. It also suppressed the formation of scar tissue while enhancing cardiac function. The data suggests that one of the beneficial effects of STC-1 might be attributed to suppression of CD14 on recruited monocytes and macrophages that limits their inflammatory response. STC-1 may be a promising therapy to protect the heart and other tissues from ischemic injury.
Copyright © 2016 Elsevier Inc. All rights reserved.
Conflict of interest statement
All authors are in agreement with the content of the manuscript and have read the journal's authorship agreement as well as the policy on disclosure of potential conflicts of interest.
Figures
References
-
- Yeung BH, Law AY, Wong CK. Evolution and roles of stanniocalcin. Mol Cell Endocrinol. 2012;349(2):272–280. - PubMed
-
- Sazonova O, James KA, McCudden CR, Segal D, Talebian A, Wagner GF. Stanniocalcin-1 secretion and receptor regulation in kidney cells. Am J Physiol Renal Physiol. 2008;294(4):F788–94. - PubMed
-
- Sheikh-Hamad D, Bick R, Wu GY, et al. Stanniocalcin-1 is a naturally occurring L-channel inhibitor in cardiomyocytes: relevance to human heart failure. Am J Physiol Heart Circ Physiol. 2003;285(1):H442–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
