

Engineering tyrosine electron transfer pathways decreases oxidative toxicity in hemoglobin: implications for blood substitute design
- PMID: 27470146
- PMCID: PMC5095908
- DOI: 10.1042/BCJ20160243
Engineering tyrosine electron transfer pathways decreases oxidative toxicity in hemoglobin: implications for blood substitute design
Abstract
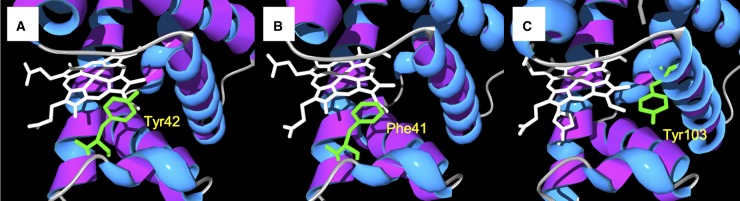
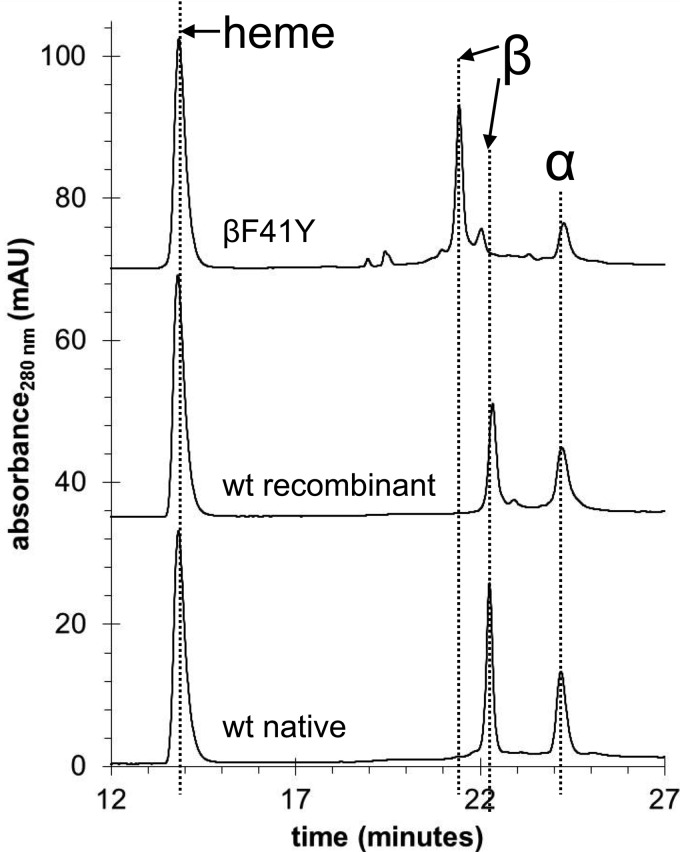
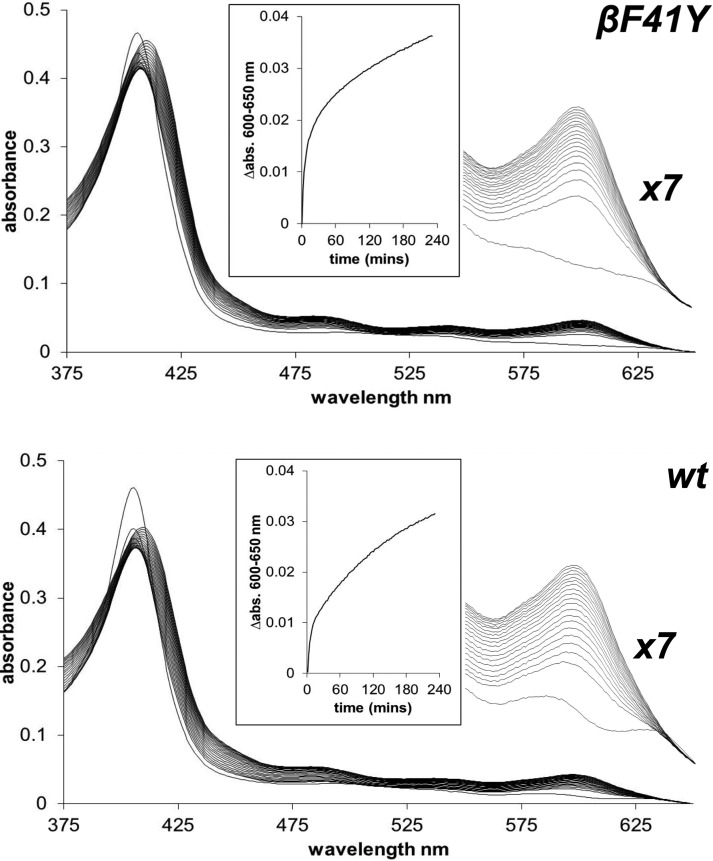
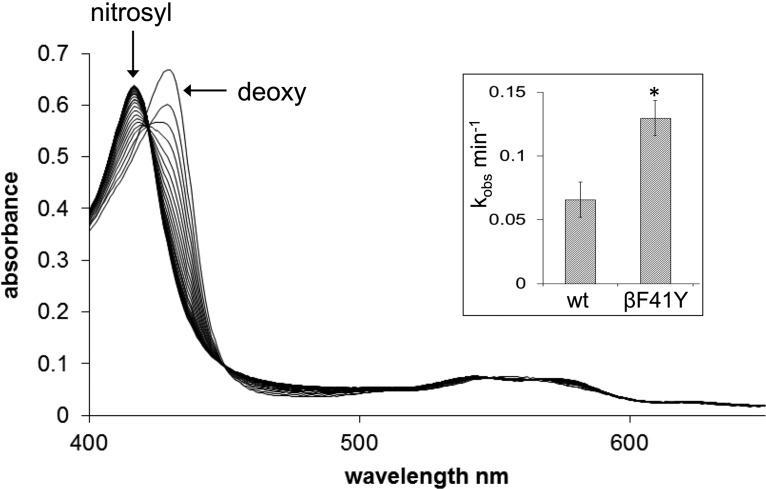
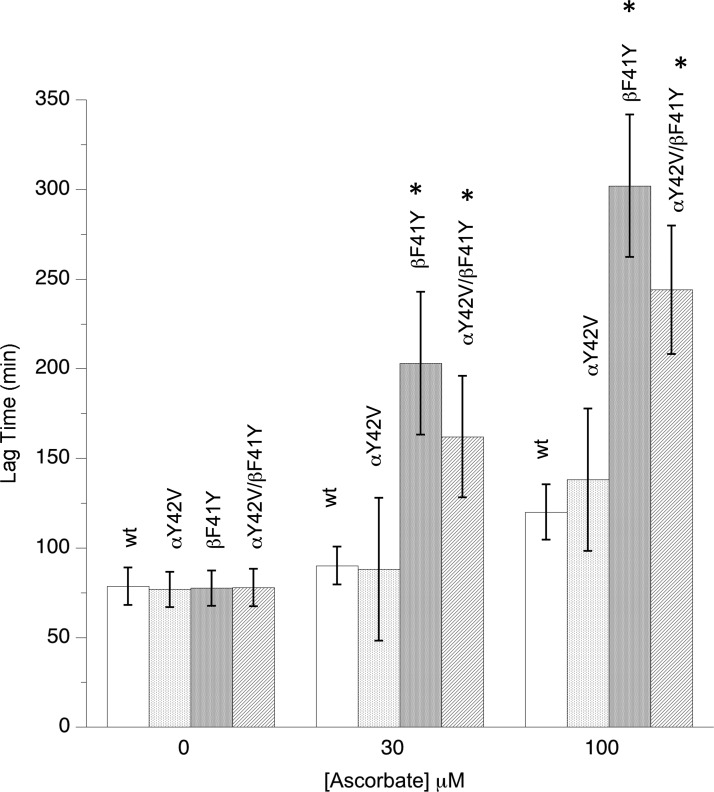
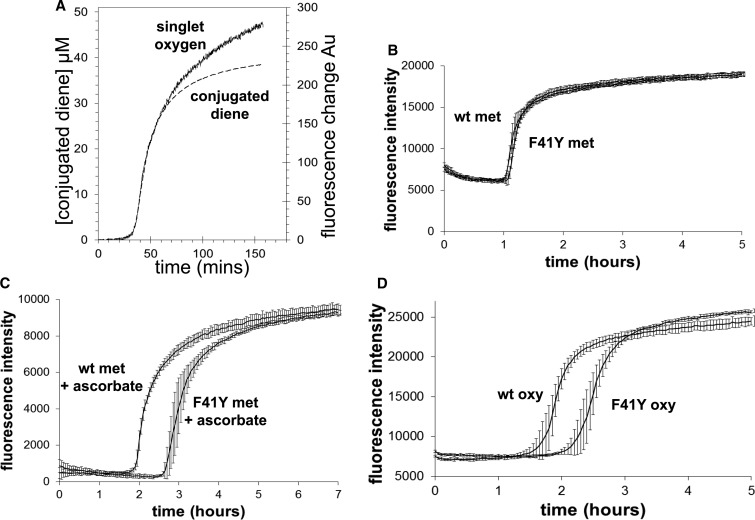
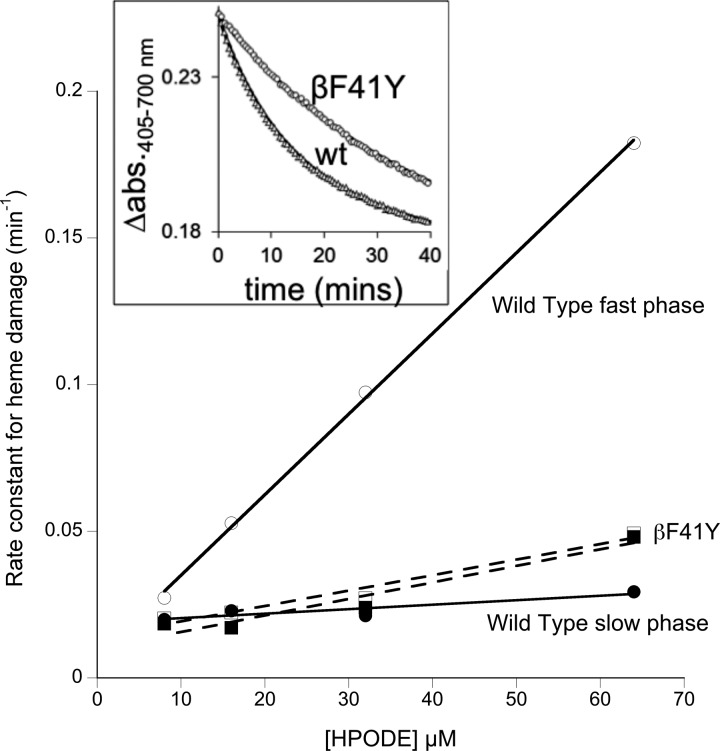
Hemoglobin (Hb)-based oxygen carriers (HBOC) have been engineered to replace or augment the oxygen-carrying capacity of erythrocytes. However, clinical results have generally been disappointing due to adverse side effects linked to intrinsic heme-mediated oxidative toxicity and nitric oxide (NO) scavenging. Redox-active tyrosine residues can facilitate electron transfer between endogenous antioxidants and oxidative ferryl heme species. A suitable residue is present in the α-subunit (Y42) of Hb, but absent from the homologous position in the β-subunit (F41). We therefore replaced this residue with a tyrosine (βF41Y, Hb Mequon). The βF41Y mutation had no effect on the intrinsic rate of lipid peroxidation as measured by conjugated diene and singlet oxygen formation following the addition of ferric(met) Hb to liposomes. However, βF41Y significantly decreased these rates in the presence of physiological levels of ascorbate. Additionally, heme damage in the β-subunit following the addition of the lipid peroxide hydroperoxyoctadecadieoic acid was five-fold slower in βF41Y. NO bioavailability was enhanced in βF41Y by a combination of a 20% decrease in NO dioxygenase activity and a doubling of the rate of nitrite reductase activity. The intrinsic rate of heme loss from methemoglobin was doubled in the β-subunit, but unchanged in the α-subunit. We conclude that the addition of a redox-active tyrosine mutation in Hb able to transfer electrons from plasma antioxidants decreases heme-mediated oxidative reactivity and enhances NO bioavailability. This class of mutations has the potential to decrease adverse side effects as one component of a HBOC product.
Keywords: hemoglobin; oxidative stress; reactive oxygen species.
© 2016 The Author(s).
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous