

Partial Activation of SA- and JA-Defensive Pathways in Strawberry upon Colletotrichum acutatum Interaction
- PMID: 27471515
- PMCID: PMC4945649
- DOI: 10.3389/fpls.2016.01036
Partial Activation of SA- and JA-Defensive Pathways in Strawberry upon Colletotrichum acutatum Interaction
Abstract
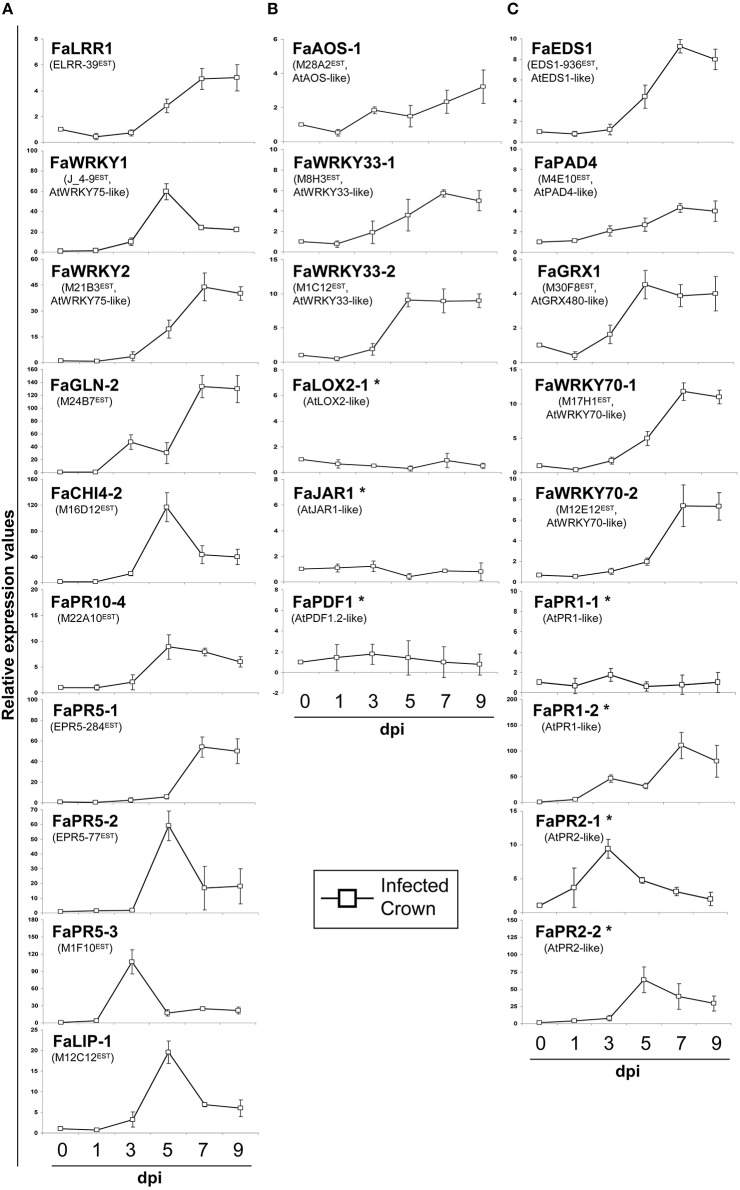
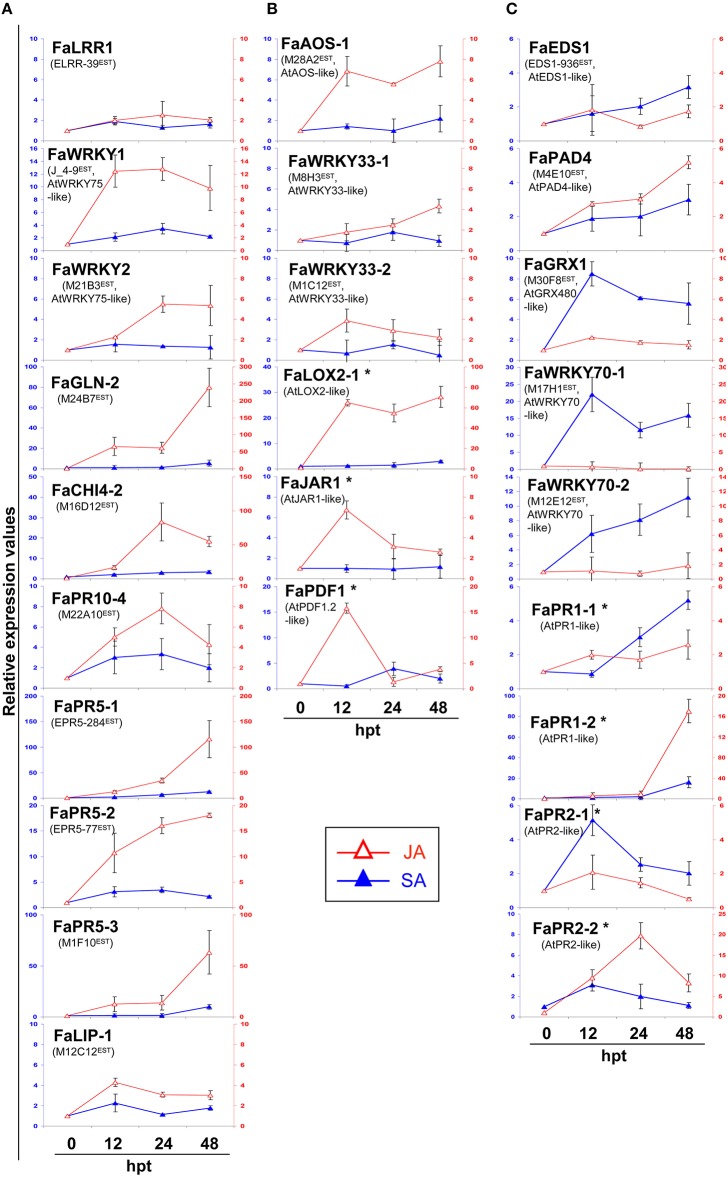
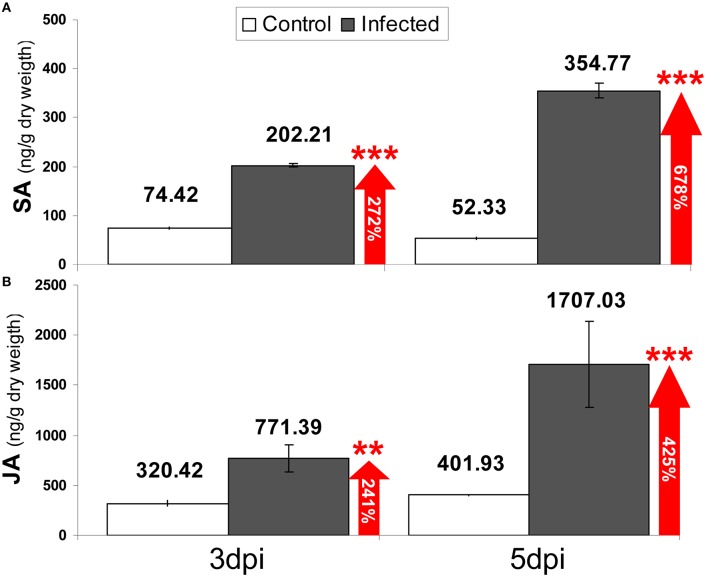
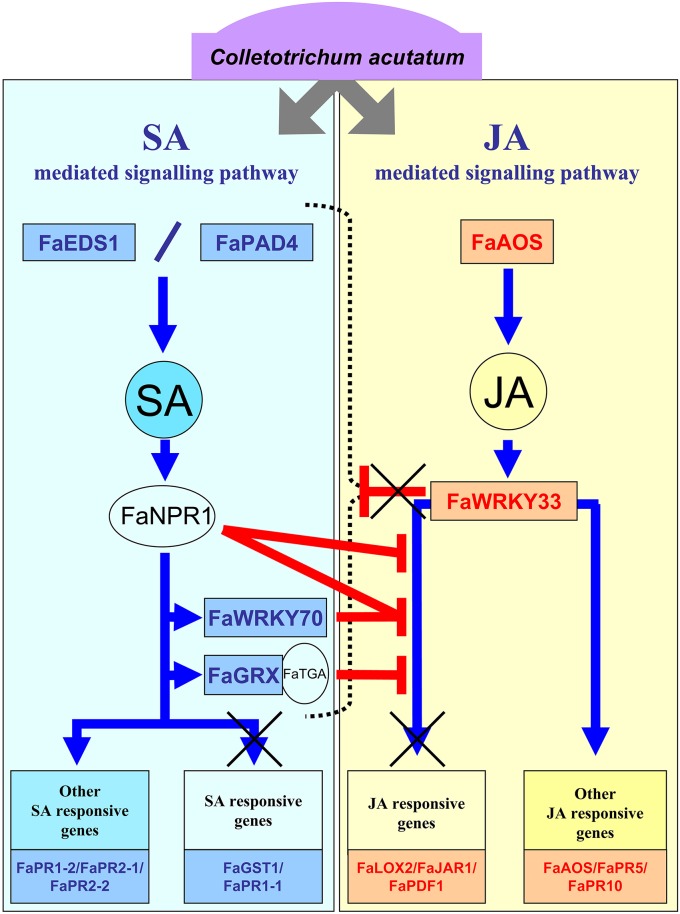
Understanding the nature of pathogen host interaction may help improve strawberry (Fragaria × ananassa) cultivars. Plant resistance to pathogenic agents usually operates through a complex network of defense mechanisms mediated by a diverse array of signaling molecules. In strawberry, resistance to a variety of pathogens has been reported to be mostly polygenic and quantitatively inherited, making it difficult to associate molecular markers with disease resistance genes. Colletotrichum acutatum spp. is a major strawberry pathogen, and completely resistant cultivars have not been reported. Moreover, strawberry defense network components and mechanisms remain largely unknown and poorly understood. Assessment of the strawberry response to C. acutatum included a global transcript analysis, and acidic hormones SA and JA measurements were analyzed after challenge with the pathogen. Induction of transcripts corresponding to the SA and JA signaling pathways and key genes controlling major steps within these defense pathways was detected. Accordingly, SA and JA accumulated in strawberry after infection. Contrastingly, induction of several important SA, JA, and oxidative stress-responsive defense genes, including FaPR1-1, FaLOX2, FaJAR1, FaPDF1, and FaGST1, was not detected, which suggests that specific branches in these defense pathways (those leading to FaPR1-2, FaPR2-1, FaPR2-2, FaAOS, FaPR5, and FaPR10) were activated. Our results reveal that specific aspects in SA and JA dependent signaling pathways are activated in strawberry upon interaction with C. acutatum. Certain described defense-associated transcripts related to these two known signaling pathways do not increase in abundance following infection. This finding suggests new insight into a specific putative molecular strategy for defense against this pathogen.
Keywords: Colletotrichum acutatum; Fragaria × ananassa; quantification of gene expression; salicylic and jasmonic acid; strawberry defense response.
Figures
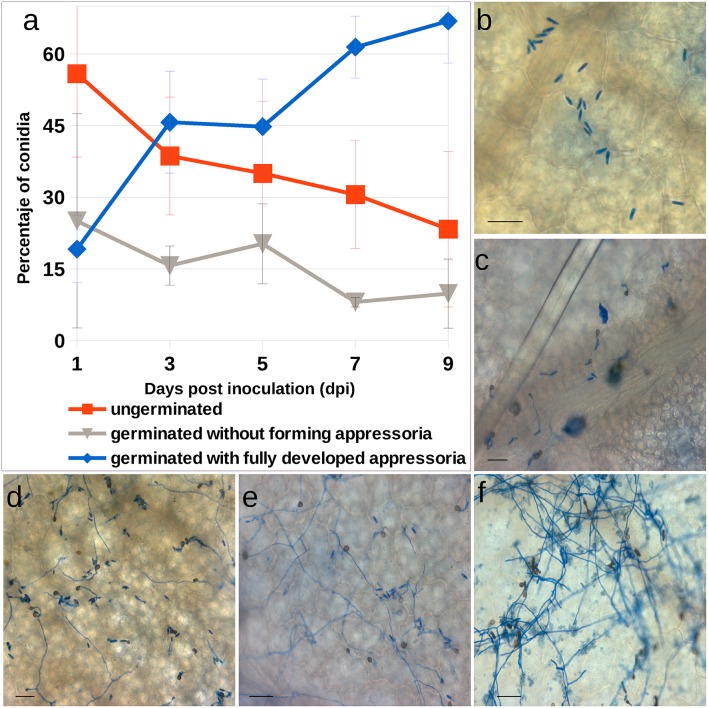
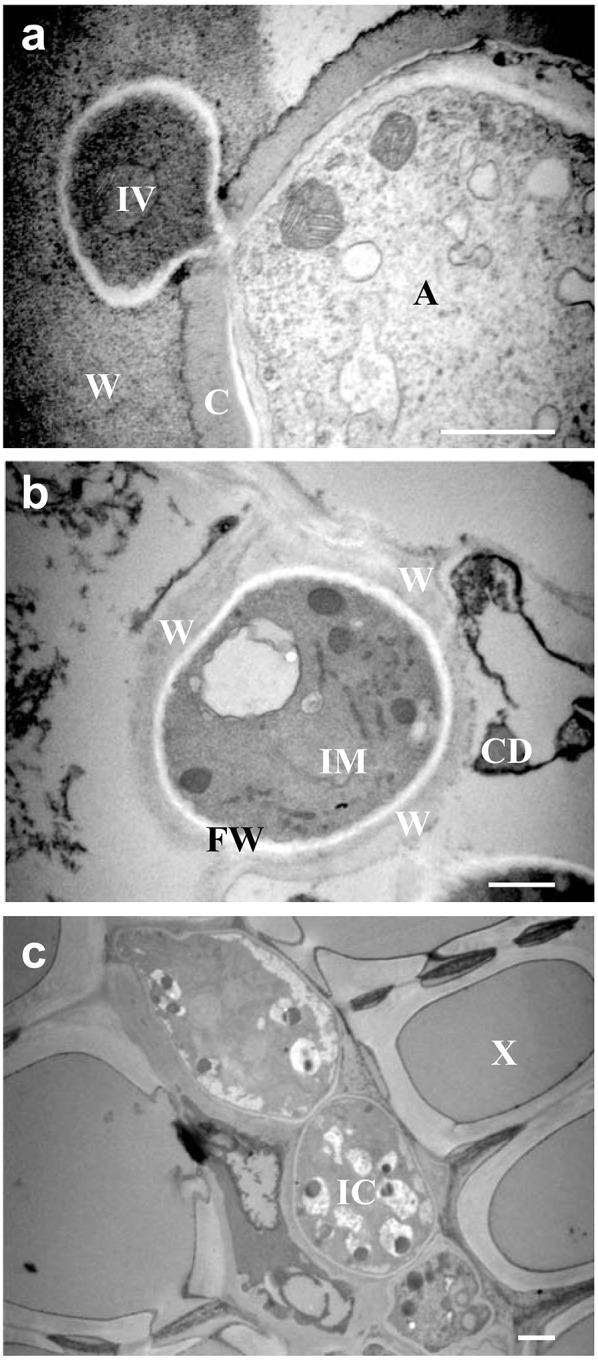
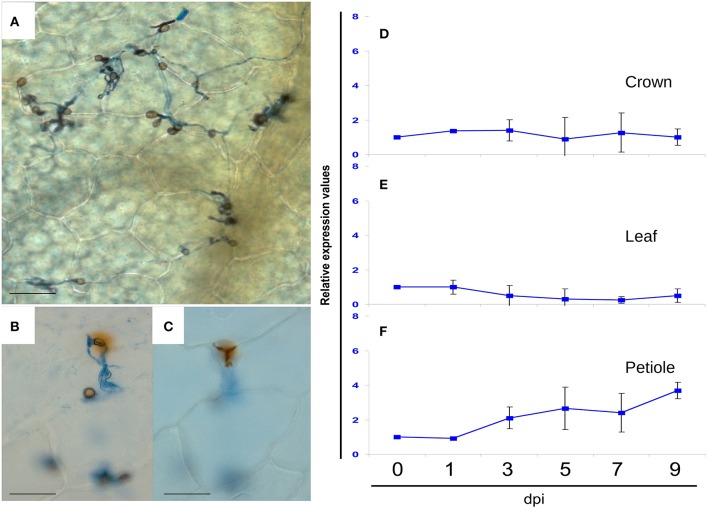
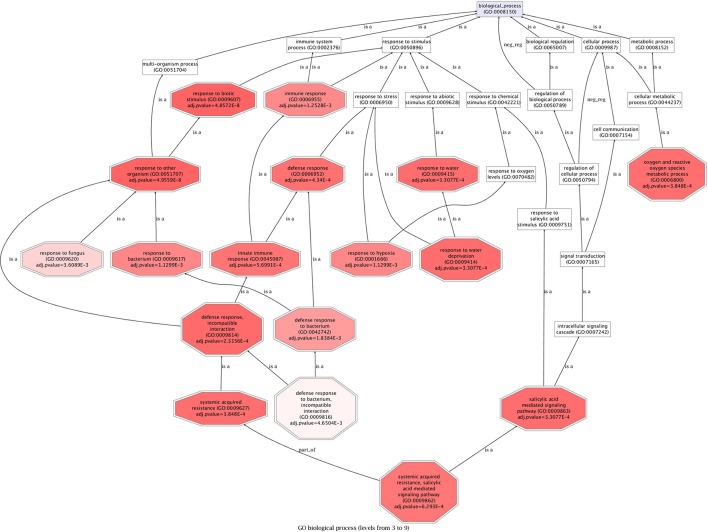
References
-
- Amil-Ruiz F., Encinas-Villarejo S., de los Santos B., Muñoz-Mérida A., Mercado J. A., Trelles O., et al. (2012). Distinctive transcriptome response of two strawberry (Fragaria x ananassa) cultivars to Colletotrichum acutatum infection, in XXVIII International Horticultural Congress on Science and Horticulture for People (IHC2010): International Symposium on Genomics and Genetic Transformation of Horticultural Crops, eds Litz R. E., Folta K., Talon M., Pliego Alfaro F. (Lisbon: ISHS; ), 47–50.
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous