

Mechanistic and Evolutionary Insights from the Reciprocal Promiscuity of Two Pyridoxal Phosphate-dependent Enzymes
- PMID: 27474741
- PMCID: PMC5025676
- DOI: 10.1074/jbc.M116.739557
Mechanistic and Evolutionary Insights from the Reciprocal Promiscuity of Two Pyridoxal Phosphate-dependent Enzymes
Abstract
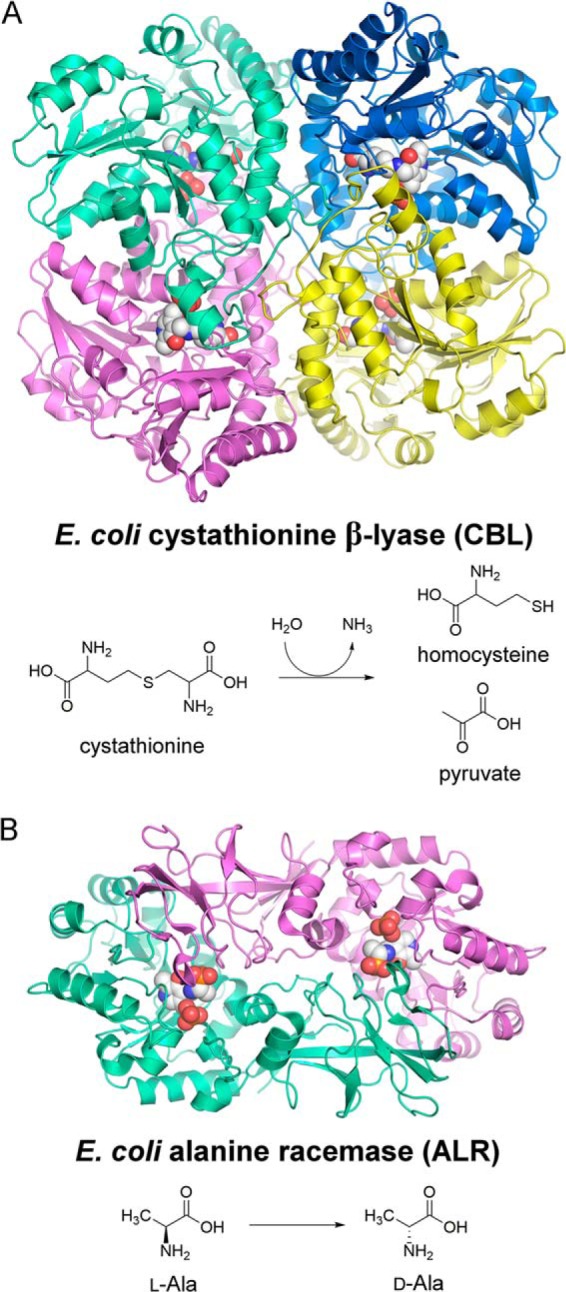
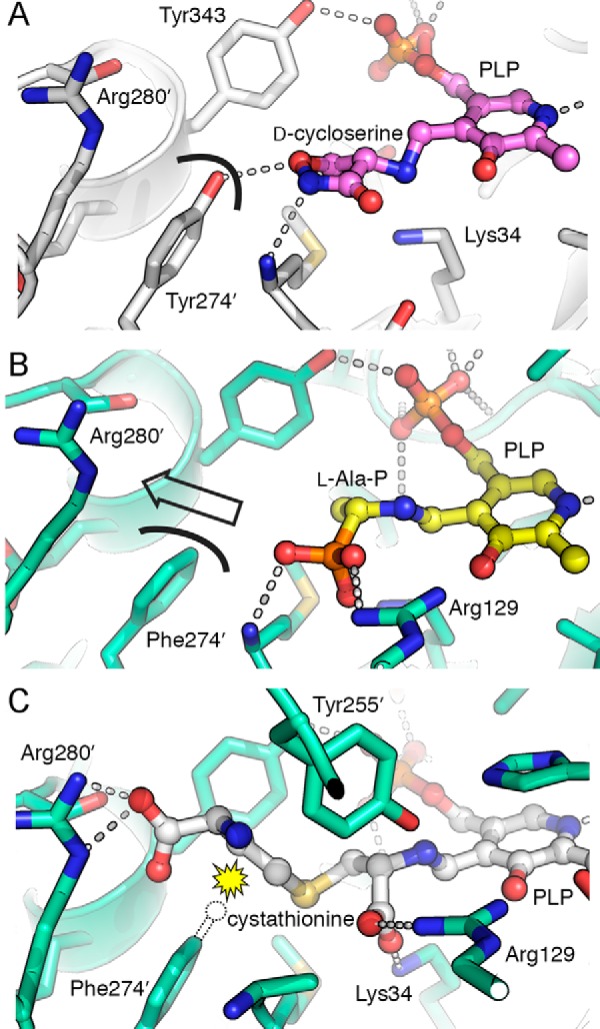
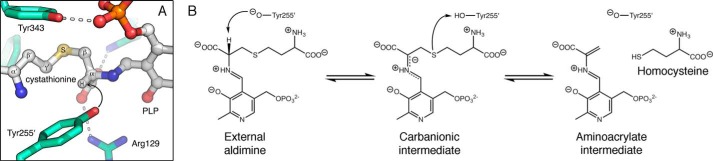
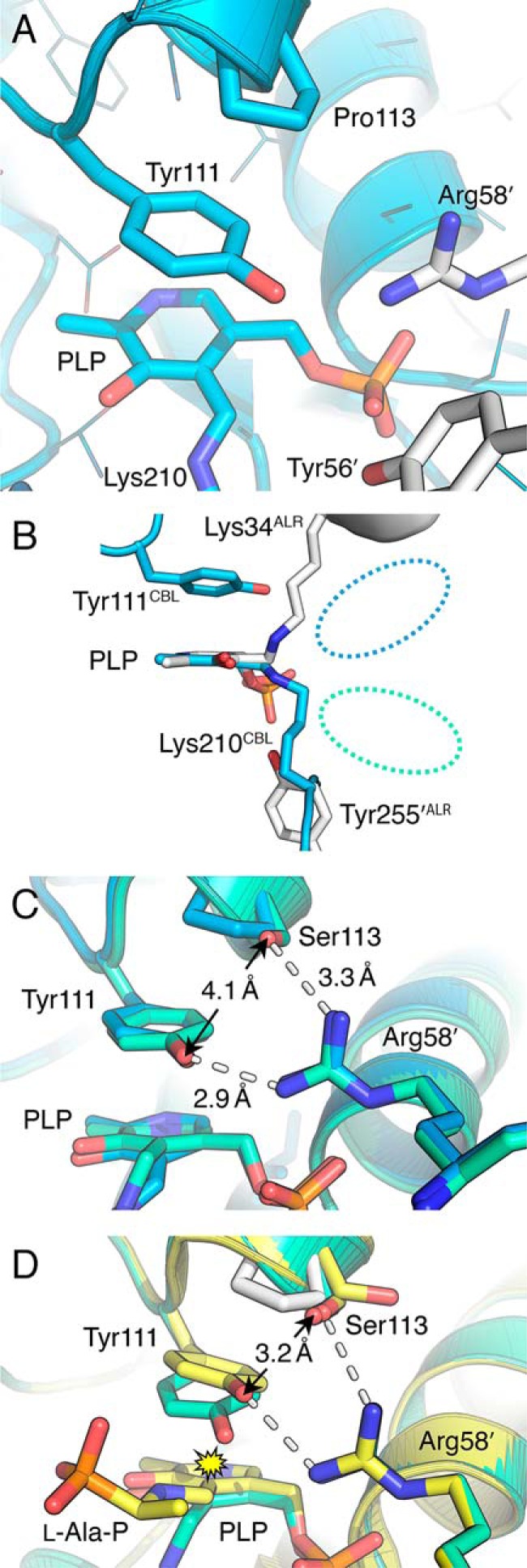
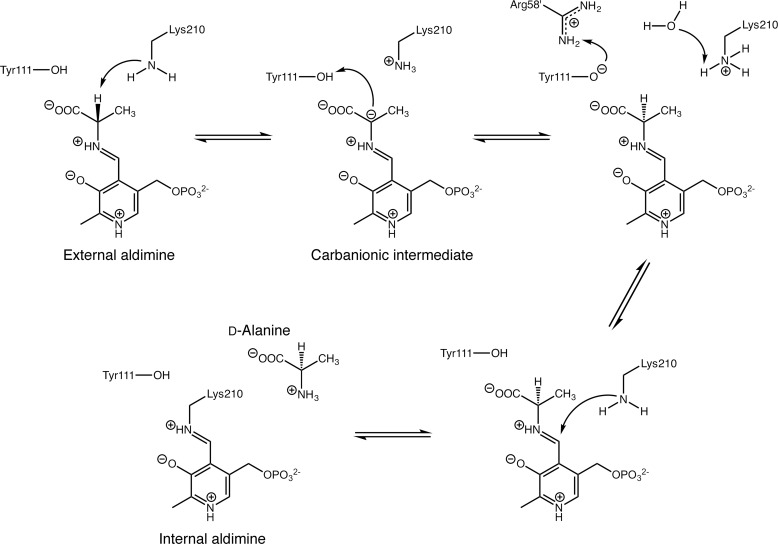
Enzymes that utilize the cofactor pyridoxal 5'-phosphate play essential roles in amino acid metabolism in all organisms. The cofactor is used by proteins that adopt at least five different folds, which raises questions about the evolutionary processes that might explain the observed distribution of functions among folds. In this study, we show that a representative of fold type III, the Escherichia coli alanine racemase (ALR), is a promiscuous cystathionine β-lyase (CBL). Furthermore, E. coli CBL (fold type I) is a promiscuous alanine racemase. A single round of error-prone PCR and selection yielded variant ALR(Y274F), which catalyzes cystathionine β-elimination with a near-native Michaelis constant (Km = 3.3 mm) but a poor turnover number (kcat ≈10 h(-1)). In contrast, directed evolution also yielded CBL(P113S), which catalyzes l-alanine racemization with a poor Km (58 mm) but a high kcat (22 s(-1)). The structures of both variants were solved in the presence and absence of the l-alanine analogue, (R)-1-aminoethylphosphonic acid. As expected, the ALR active site was enlarged by the Y274F substitution, allowing better access for cystathionine. More surprisingly, the favorable kinetic parameters of CBL(P113S) appear to result from optimizing the pKa of Tyr-111, which acts as the catalytic acid during l-alanine racemization. Our data emphasize the short mutational routes between the functions of pyridoxal 5'-phosphate-dependent enzymes, regardless of whether or not they share the same fold. Thus, they confound the prevailing model of enzyme evolution, which predicts that overlapping patterns of promiscuity result from sharing a common multifunctional ancestor.
Keywords: directed evolution; enzyme mechanism; protein evolution; protein structure; pyridoxal phosphate.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Raboni S., Spyrakis F., Campanini B., Amadasi A., Bettati S., Peracchi A., Mozzarelli A., and Contestabile R. (2010) in Comprehensive Natural Products II: Chemistry and Biology (Mander L. and Liu H.-W., editors in chief) pp. 273–350, Elsevier Ltd., New York
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
