

Accumulation of Peptidoglycan O-Acetylation Leads to Altered Cell Wall Biochemistry and Negatively Impacts Pathogenesis Factors of Campylobacter jejuni
- PMID: 27474744
- PMCID: PMC5077204
- DOI: 10.1074/jbc.M116.746404
Accumulation of Peptidoglycan O-Acetylation Leads to Altered Cell Wall Biochemistry and Negatively Impacts Pathogenesis Factors of Campylobacter jejuni
Abstract
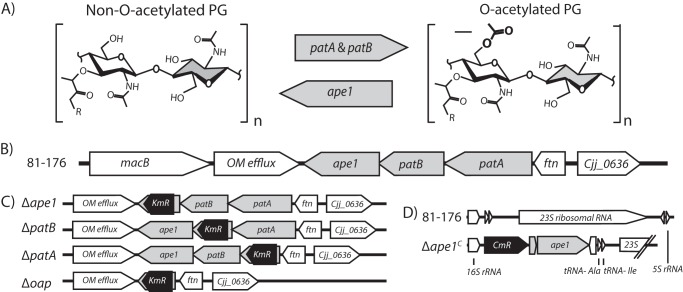
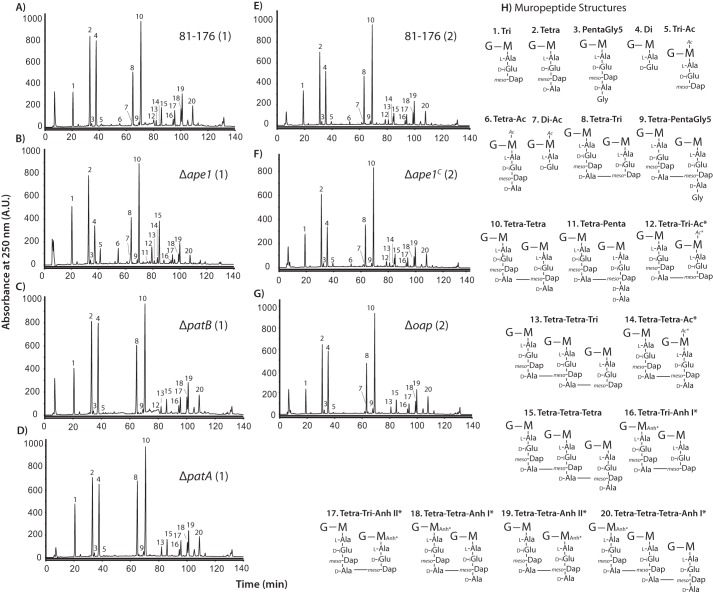
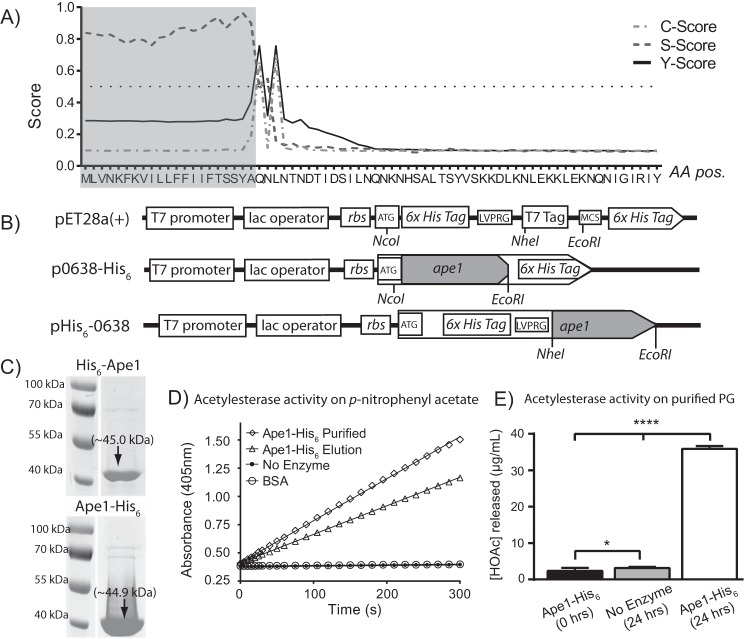
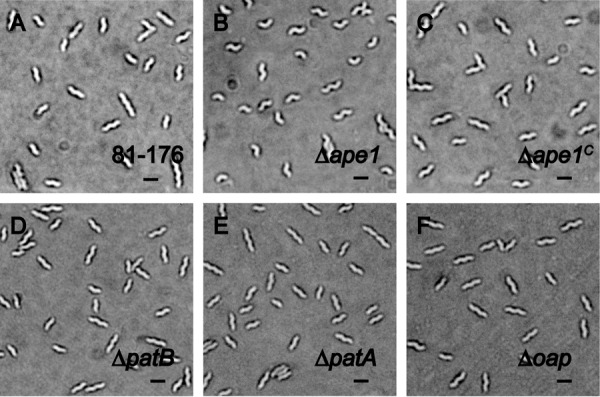
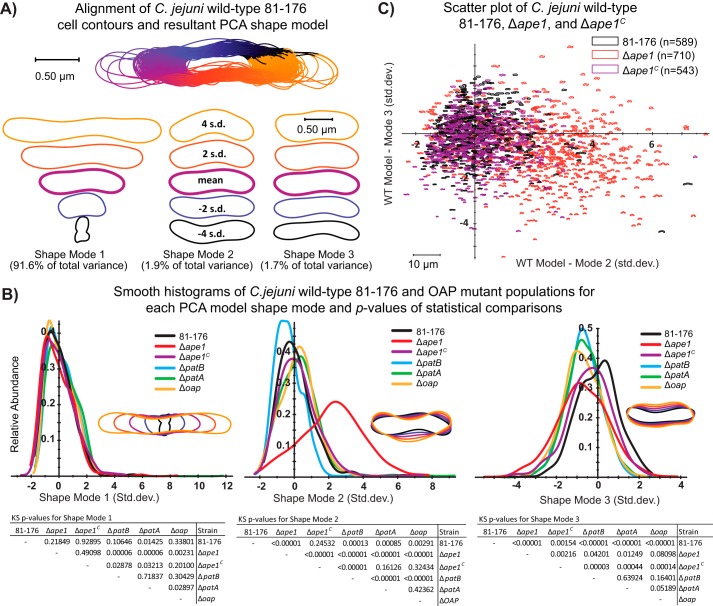
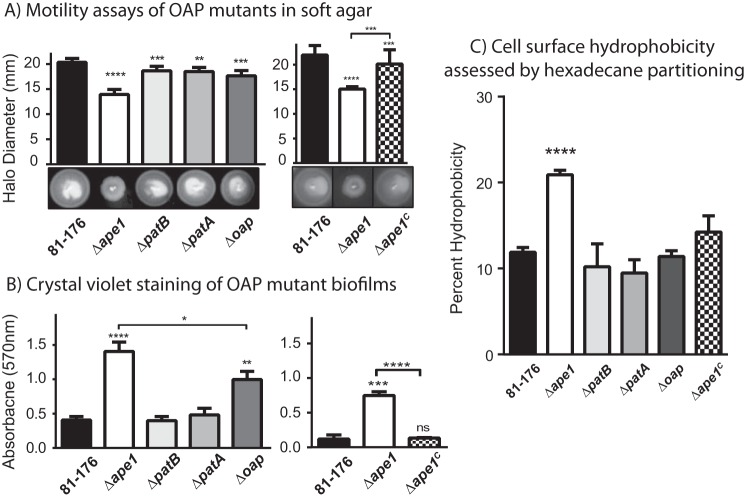
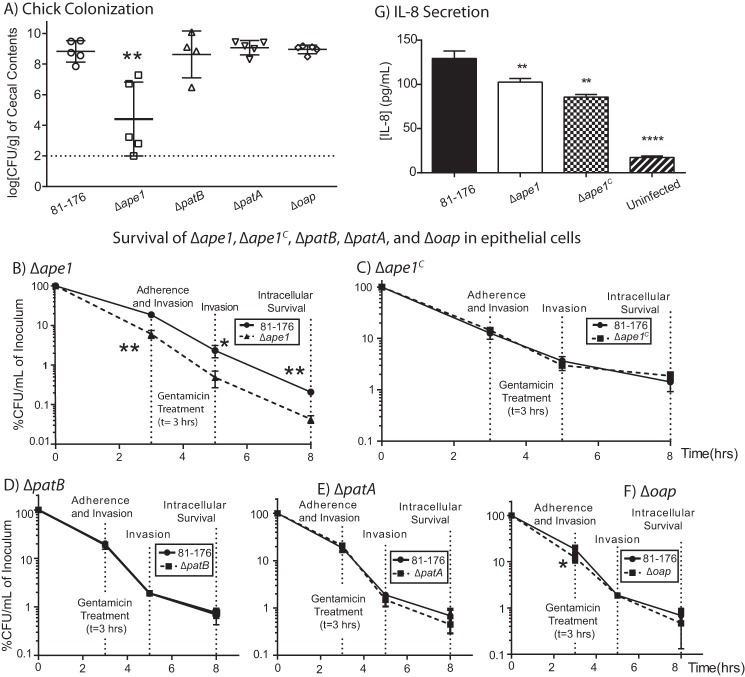
Campylobacter jejuni is a leading cause of bacterial gastroenteritis in the developed world. Despite its prevalence, its mechanisms of pathogenesis are poorly understood. Peptidoglycan (PG) is important for helical shape, colonization, and host-pathogen interactions in C. jejuni Therefore, changes in PG greatly impact the physiology of this organism. O-acetylation of peptidoglycan (OAP) is a bacterial phenomenon proposed to be important for proper cell growth, characterized by acetylation of the C6 hydroxyl group of N-acetylmuramic acid in the PG glycan backbone. The OAP gene cluster consists of a PG O-acetyltransferase A (patA) for translocation of acetate into the periplasm, a PG O-acetyltransferase B (patB) for O-acetylation, and an O-acetylpeptidoglycan esterase (ape1) for de-O-acetylation. In this study, reduced OAP in ΔpatA and ΔpatB had minimal impact on C. jejuni growth and fitness under the conditions tested. However, accumulation of OAP in Δape1 resulted in marked differences in PG biochemistry, including O-acetylation, anhydromuropeptide levels, and changes not expected to result directly from Ape1 activity. This suggests that OAP may be a form of substrate level regulation in PG biosynthesis. Ape1 acetylesterase activity was confirmed in vitro using p-nitrophenyl acetate and O-acetylated PG as substrates. In addition, Δape1 exhibited defects in pathogenesis-associated phenotypes, including cell shape, motility, biofilm formation, cell surface hydrophobicity, and sodium deoxycholate sensitivity. Δape1 was also impaired for chick colonization and adhesion, invasion, intracellular survival, and induction of IL-8 production in INT407 cells in vitro The importance of Ape1 in C. jejuni biology makes it a good candidate as an antimicrobial target.
Keywords: Campylobacter jejuni; O-Acetylation; O-Acetylesterase; acetylation; bacterial pathogenesis; cell wall; gram-negative bacteria; peptidoglycan.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
Similar articles
-
Peptidoglycan Acetylation of Campylobacter jejuni Is Essential for Maintaining Cell Wall Integrity and Colonization in Chicken Intestines.Appl Environ Microbiol. 2016 Sep 30;82(20):6284-6290. doi: 10.1128/AEM.02068-16. Print 2016 Oct 15. Appl Environ Microbiol. 2016. PMID: 27520822 Free PMC article.
-
Assay for peptidoglycan O-acetyltransferase: a potential new antibacterial target.Anal Biochem. 2013 Aug 15;439(2):73-9. doi: 10.1016/j.ab.2013.04.022. Epub 2013 May 7. Anal Biochem. 2013. PMID: 23660013
-
The Campylobacter jejuni helical to coccoid transition involves changes to peptidoglycan and the ability to elicit an immune response.Mol Microbiol. 2019 Jul;112(1):280-301. doi: 10.1111/mmi.14269. Epub 2019 May 20. Mol Microbiol. 2019. PMID: 31070821 Free PMC article.
-
Campylobacter jejuni: targeting host cells, adhesion, invasion, and survival.Appl Microbiol Biotechnol. 2023 May;107(9):2725-2754. doi: 10.1007/s00253-023-12456-w. Epub 2023 Mar 21. Appl Microbiol Biotechnol. 2023. PMID: 36941439 Free PMC article. Review.
-
Mechanistic Pathways for Peptidoglycan O-Acetylation and De-O-Acetylation.Front Microbiol. 2018 Oct 1;9:2332. doi: 10.3389/fmicb.2018.02332. eCollection 2018. Front Microbiol. 2018. PMID: 30327644 Free PMC article. Review.
Cited by
-
Hyperbiofilm Formation by Bordetella pertussis Strains Correlates with Enhanced Virulence Traits.Infect Immun. 2017 Nov 17;85(12):e00373-17. doi: 10.1128/IAI.00373-17. Print 2017 Dec. Infect Immun. 2017. PMID: 28893915 Free PMC article.
-
Mechanism of D-alanine transfer to teichoic acids shows how bacteria acylate cell envelope polymers.Nat Microbiol. 2023 Jul;8(7):1318-1329. doi: 10.1038/s41564-023-01411-0. Epub 2023 Jun 12. Nat Microbiol. 2023. PMID: 37308592 Free PMC article.
-
Streptococcus pneumoniae synchronizes the states of cell wall peptidoglycan acetylation and genome methylation by programmed DNA inversions.PLoS Pathog. 2025 Aug 5;21(8):e1013286. doi: 10.1371/journal.ppat.1013286. eCollection 2025 Aug. PLoS Pathog. 2025. PMID: 40763171 Free PMC article.
-
Multiple Campylobacter jejuni proteins affecting the peptidoglycan structure and the degree of helical cell curvature.Front Microbiol. 2023 Apr 18;14:1162806. doi: 10.3389/fmicb.2023.1162806. eCollection 2023. Front Microbiol. 2023. PMID: 37143542 Free PMC article.
-
Interplay between Peptidoglycan Biology and Virulence in Gram-Negative Pathogens.Microbiol Mol Biol Rev. 2018 Sep 12;82(4):e00033-18. doi: 10.1128/MMBR.00033-18. Print 2018 Dec. Microbiol Mol Biol Rev. 2018. PMID: 30209071 Free PMC article. Review.
References
-
- Yuki N., and Hartung H. P. (2012) Guillain-Barre syndrome. N. Engl. J. Med. 366, 2294–2304 - PubMed
-
- Jacobs B. C., Rothbarth P. H., van der Meché F. G., Herbrink P., Schmitz P. I., de Klerk M. A., and van Doorn P. A. (1998) The spectrum of antecedent infections in Guillain-Barre syndrome: a case-control study. Neurology 51, 1110–1115 - PubMed
-
- Man S. M. (2011) The clinical importance of emerging Campylobacter species. Nat. Rev. Gastroenterol. Hepatol. 8, 669–685 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
