

Extracellular vesicles carry microRNA-195 to intrahepatic cholangiocarcinoma and improve survival in a rat model
- PMID: 27474881
- PMCID: PMC5258762
- DOI: 10.1002/hep.28735
Extracellular vesicles carry microRNA-195 to intrahepatic cholangiocarcinoma and improve survival in a rat model
Abstract
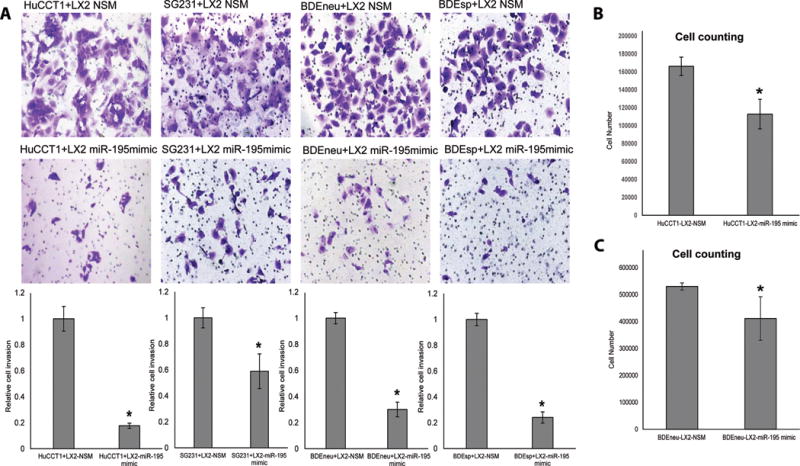
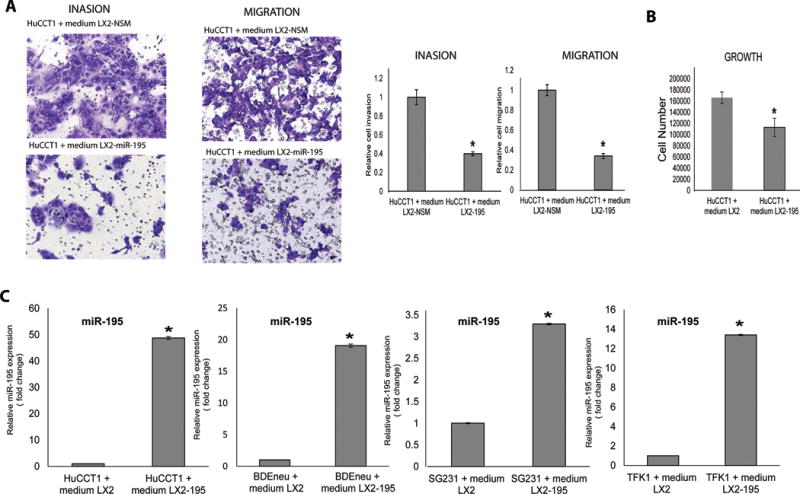
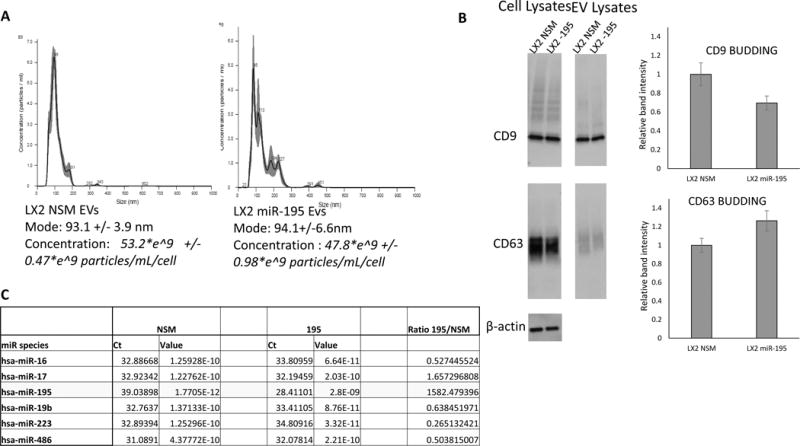
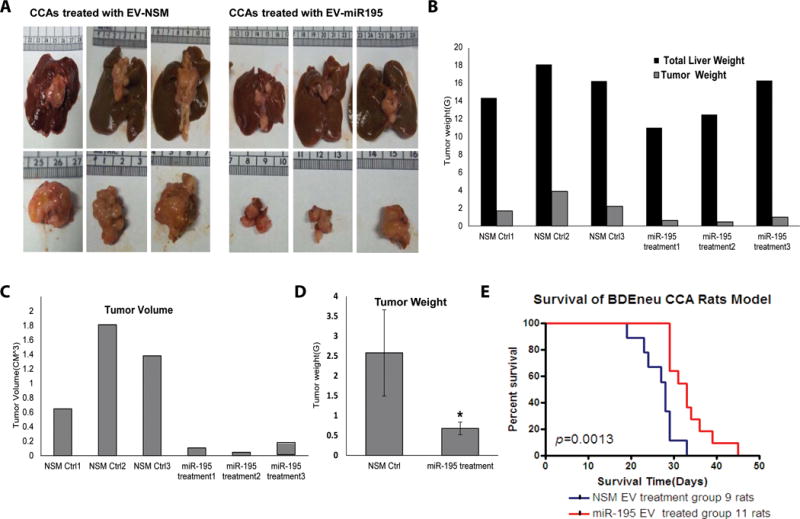
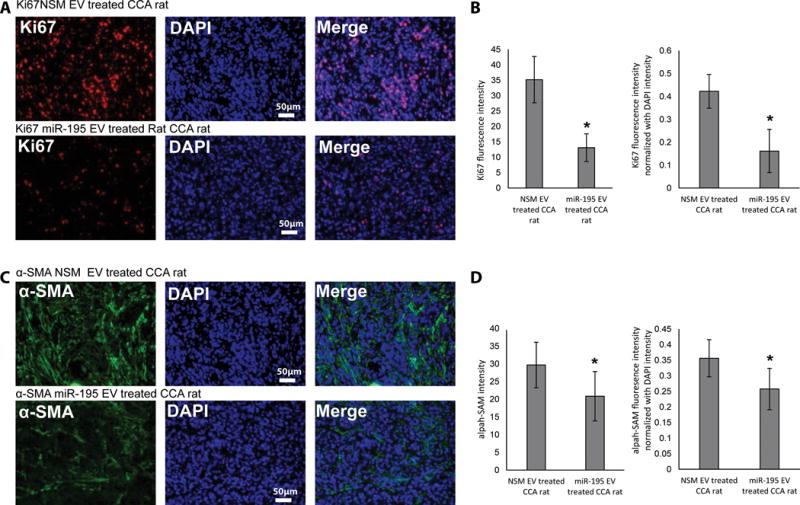
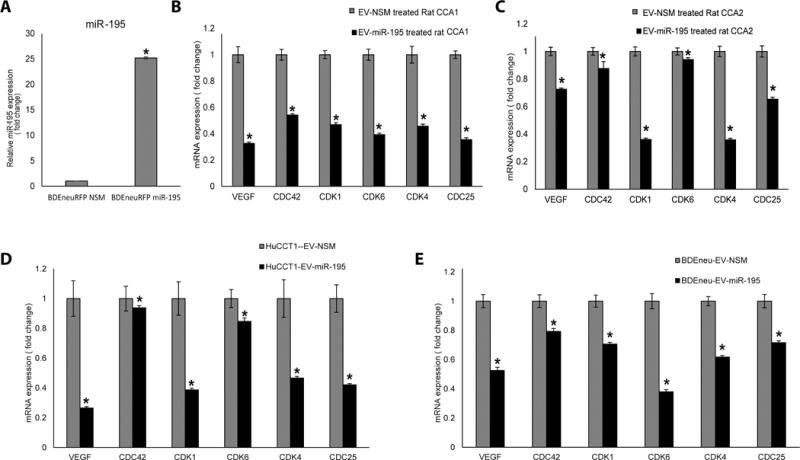
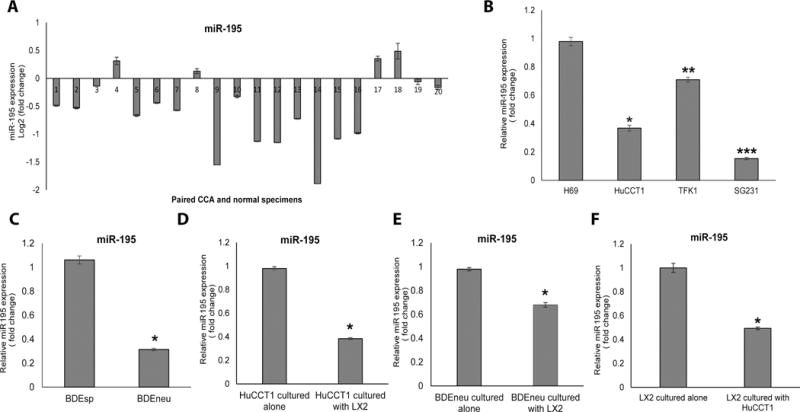
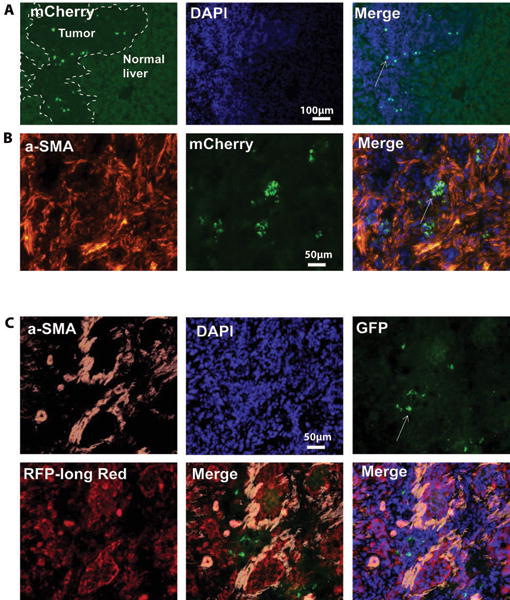
The cancer microenvironment plays a central role in cancer development, growth, and homeostasis. This paradigm suggests that cancer fibroblasts support cancers, probably in response to stimuli received from the cancer cells. We aimed at investigating whether extracellular vesicles (EVs) can shuttle microRNA (miR) species between cancer-associated fibroblasts (CAFs) and cancer cells. To this end, we extracted EVs according to published protocols. EVs were studied for their miR content by quantitative reverse-transcription polymerase chain reaction. EVs were transfected with select miR species and utilized in vitro as well as in vivo in a rat model of cholangiocarcinoma (CCA). We found that miR-195 is down-regulated in CCA cells, as well as in adjoining fibroblasts. Furthermore, we report that EVs shuttle miR-195 from fibroblasts to cancer cells. Last, we show that fibroblast-derived EVs, loaded with miR-195, can be administered in a rat model of CCA, concentrate within the tumor, decrease the size of cancers, and improve survival of treated rats.
Conclusion: EVs play a salient role in trafficking miR species between cancer cells and CAFs in human CCA. Understanding of these mechanisms may allow devising of novel therapeutics. (Hepatology 2017;65:501-514).
© 2016 by the American Association for the Study of Liver Diseases.
Conflict of interest statement
No authors have any potential conflicts in reference to this manuscript.
Figures
Comment in
-
Extracellular vesicles as therapeutic carriers of microRNAs for cholangiocarcinoma.Hepatology. 2017 Feb;65(2):404-406. doi: 10.1002/hep.28925. Epub 2016 Dec 24. Hepatology. 2017. PMID: 28012192 Free PMC article. No abstract available.
References
-
- Mueller MM, Fusenig NE. Friends or foes - bipolar effects of the tumour stroma in cancer. Nat Rev Cancer. 2004;4:839–849. - PubMed
-
- Webber JP, Spary LK, Sanders AJ, Chowdhury R, Jiang WG, Steadman R, Wymant J, et al. Differentiation of tumour-promoting stromal myofibroblasts by cancer exosomes. Oncogene. 2015;34:290–302. - PubMed
-
- Josson S, Gururajan M, Sung SY, Hu P, Shao C, Zhau HE, Liu C, et al. Stromal fibroblast-derived miR-409 promotes epithelial-to-mesenchymal transition and prostate tumorigenesis. Oncogene. 2015;34:2690–2699. - PubMed
-
- Madar S, Goldstein I, Rotter V. ‘Cancer associated fibroblasts’–more than meets the eye. Trends Mol Med. 2013;19:447–453. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
