

Polarity in plant asymmetric cell division: Division orientation and cell fate differentiation
- PMID: 27475487
- PMCID: PMC5522747
- DOI: 10.1016/j.ydbio.2016.07.020
Polarity in plant asymmetric cell division: Division orientation and cell fate differentiation
Abstract
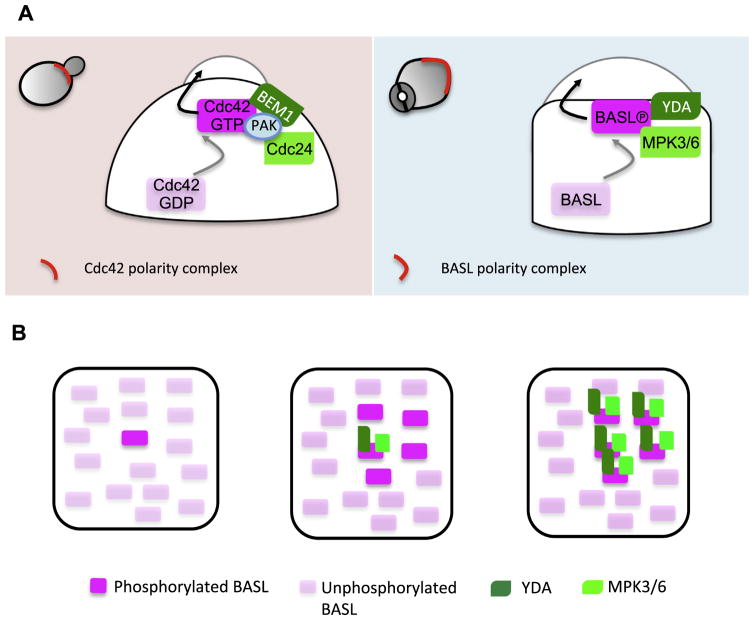
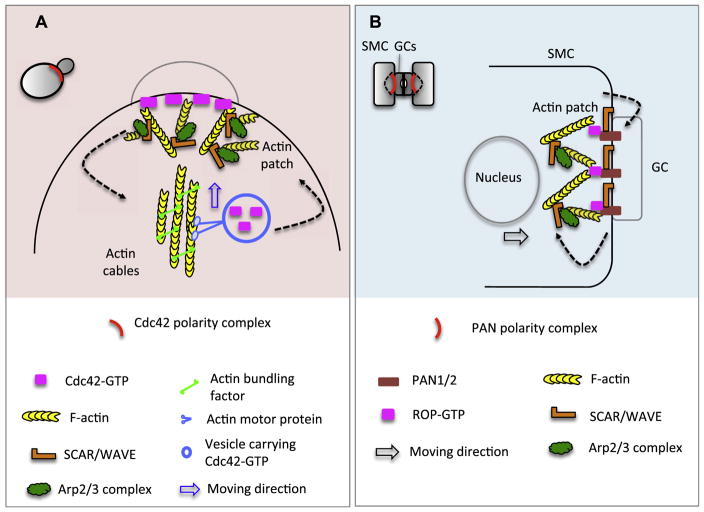
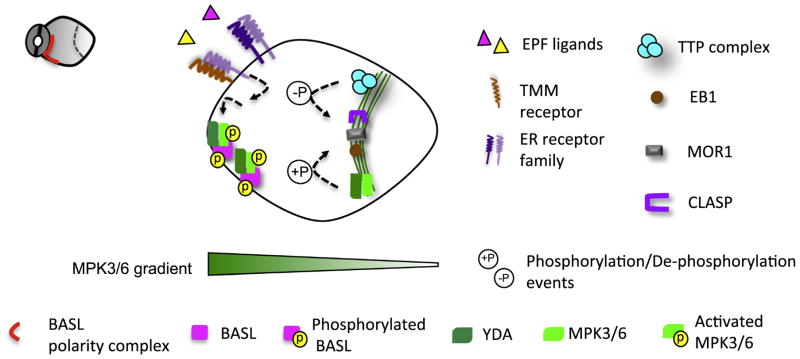
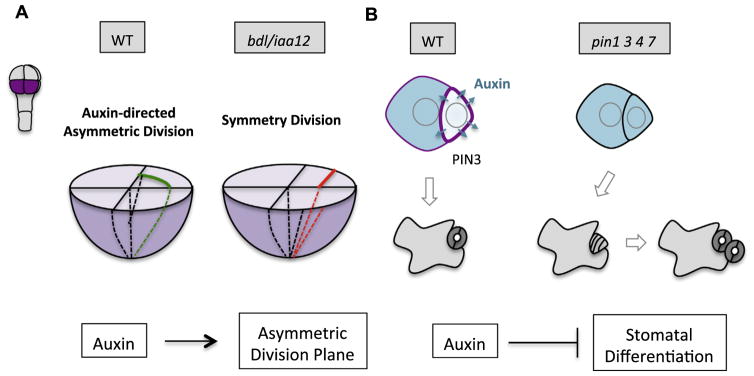
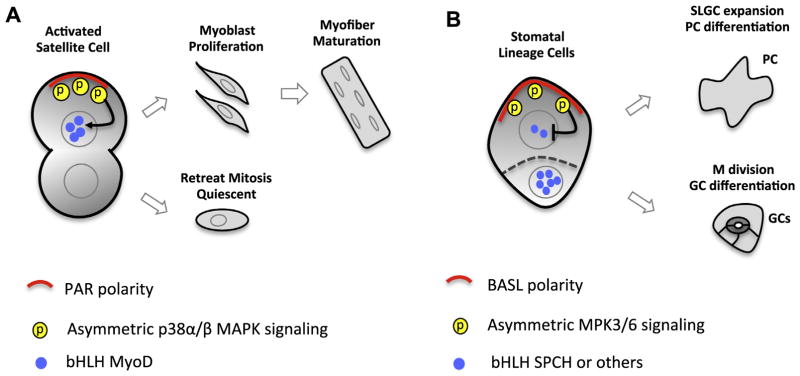
Asymmetric cell division (ACD) is universally required for the development of multicellular organisms. Unlike animal cells, plant cells have a rigid cellulosic extracellular matrix, the cell wall, which provides physical support and forms communication routes. This fundamental difference leads to some unique mechanisms in plants for generating asymmetries during cell division. However, plants also utilize intrinsically polarized proteins to regulate asymmetric signaling and cell division, a strategy similar to the differentiation mechanism found in animals. Current progress suggests that common regulatory modes, i.e. protein spontaneous clustering and cytoskeleton reorganization, underlie protein polarization in both animal and plant cells. Despite these commonalities, it is important to note that intrinsic mechanisms in plants are heavily influenced by extrinsic cues. To control physical asymmetry in cell division, although our understanding is fragmentary thus far, plants might have evolved novel polarization strategies to orientate cell division plane. Recent studies also suggest that the phytohormone auxin, one of the most pivotal small molecules in plant development, regulates ACD in plants.
Keywords: Asymmetric cell division; Auxin signaling; Cell division orientation; Cell polarity; Differential cell identity; Plant development; Protein polarization.
Copyright © 2016. Published by Elsevier Inc.
Conflict of interest statement
The authors declare that there are no conflicts of interest.
Figures
References
-
- Abrash EB, Bergmann DC. Asymmetric cell divisions: a view from plant development. Dev Cell. 2009;16:783–796. - PubMed
-
- Abrash EB, Bergmann DC. Regional specification of stomatal production by the putative ligand CHALLAH. Development. 2010;137:447–455. - PubMed
-
- Ayaydin F, Vissi E, Meszaros T, Miskolczi P, Kovacs I, Feher A, Dombradi V, Erdodi F, Gergely P, Dudits D. Inhibition of serine/threonine-specific protein phosphatases causes premature activation of cdc2MsF kinase at G2/M transition and early mitotic microtubule organisation in alfalfa. Plant J: Cell Mol Biol. 2000;23:85–96. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
