

DNA Breaks and End Resection Measured Genome-wide by End Sequencing
- PMID: 27477910
- PMCID: PMC6299834
- DOI: 10.1016/j.molcel.2016.06.034
DNA Breaks and End Resection Measured Genome-wide by End Sequencing
Abstract
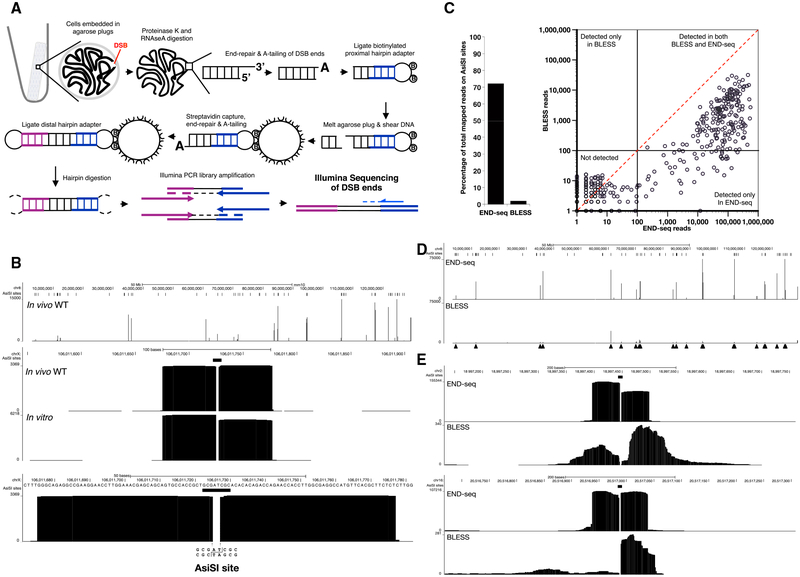
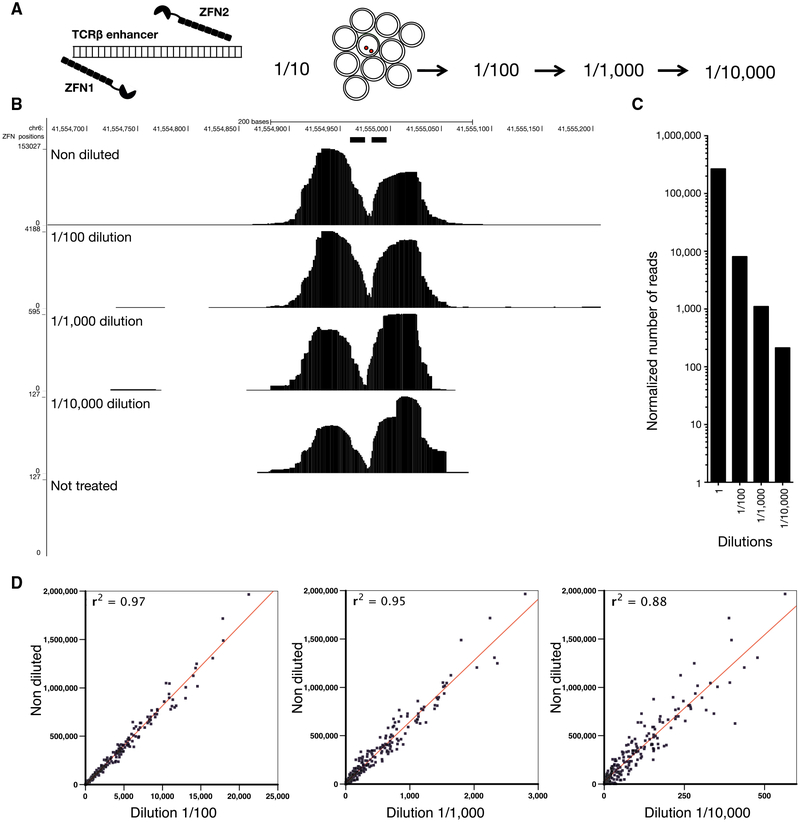
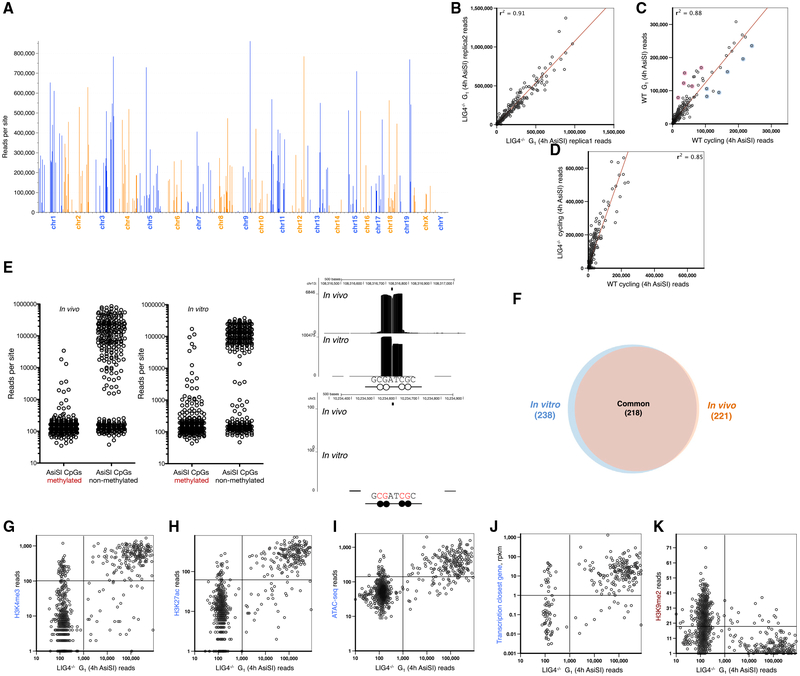
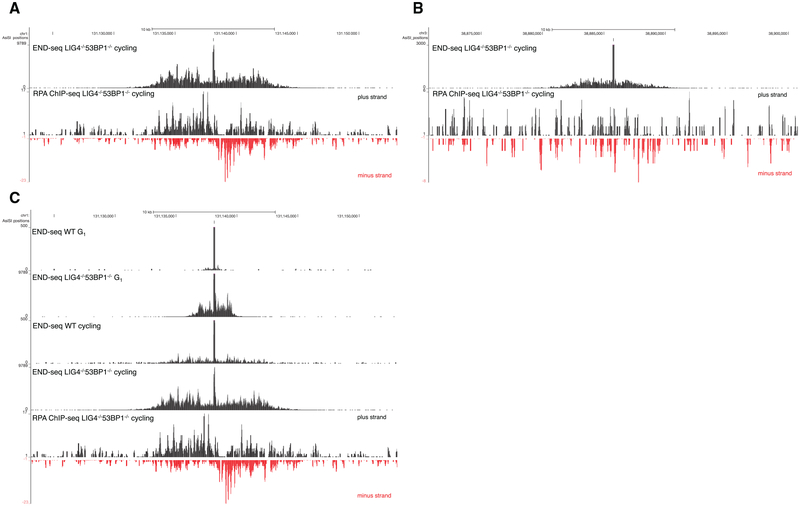
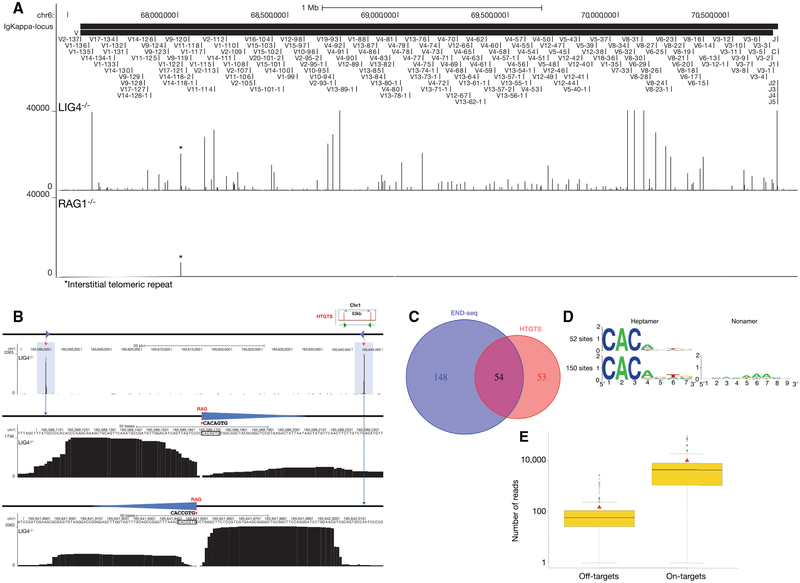
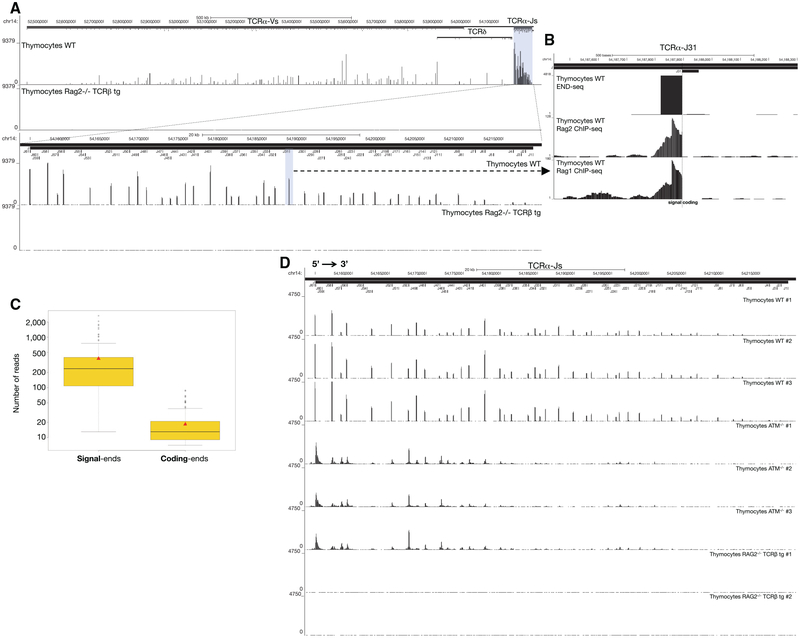
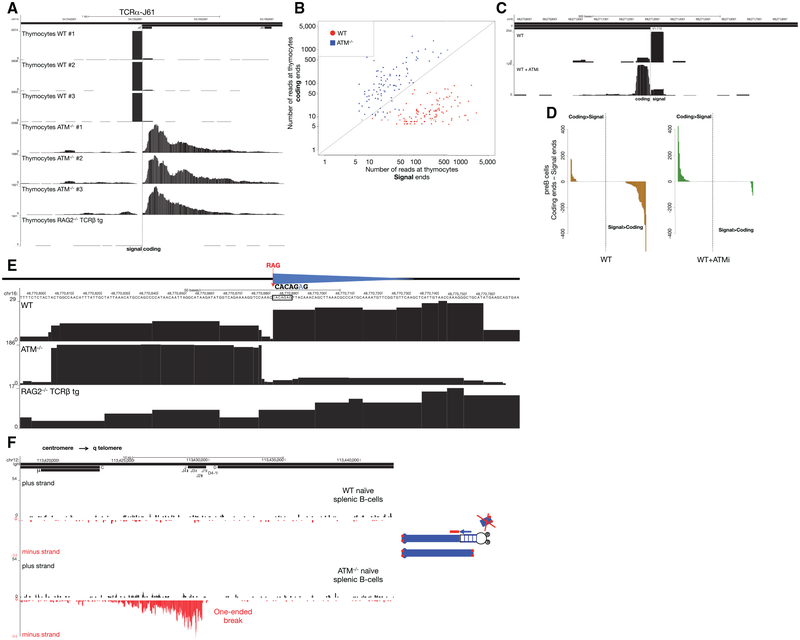
DNA double-strand breaks (DSBs) arise during physiological transcription, DNA replication, and antigen receptor diversification. Mistargeting or misprocessing of DSBs can result in pathological structural variation and mutation. Here we describe a sensitive method (END-seq) to monitor DNA end resection and DSBs genome-wide at base-pair resolution in vivo. We utilized END-seq to determine the frequency and spectrum of restriction-enzyme-, zinc-finger-nuclease-, and RAG-induced DSBs. Beyond sequence preference, chromatin features dictate the repertoire of these genome-modifying enzymes. END-seq can detect at least one DSB per cell among 10,000 cells not harboring DSBs, and we estimate that up to one out of 60 cells contains off-target RAG cleavage. In addition to site-specific cleavage, we detect DSBs distributed over extended regions during immunoglobulin class-switch recombination. Thus, END-seq provides a snapshot of DNA ends genome-wide, which can be utilized for understanding genome-editing specificities and the influence of chromatin on DSB pathway choice.
Published by Elsevier Inc.
Figures
Comment in
-
Finding DNA Ends within a Haystack of Chromatin.Mol Cell. 2016 Sep 1;63(5):726-8. doi: 10.1016/j.molcel.2016.08.012. Mol Cell. 2016. PMID: 27588600
References
-
- Bredemeyer AL, Sharma GG, Huang CY, Helmink BA, Walker LM, Khor KC, Nuskey B, Sullivan KE, Pandita TK, Bassing CH, and Sleckman BP (2006). ATM stabilizes DNA double-strand-break complexes during V(D)J recombination. Nature 442, 466–470. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
