

Early gut colonizers shape parasite susceptibility and microbiota composition in honey bee workers
- PMID: 27482088
- PMCID: PMC4995961
- DOI: 10.1073/pnas.1606631113
Early gut colonizers shape parasite susceptibility and microbiota composition in honey bee workers
Abstract
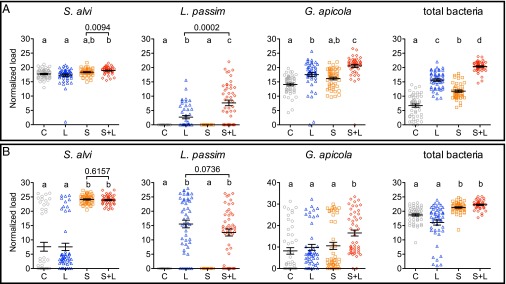
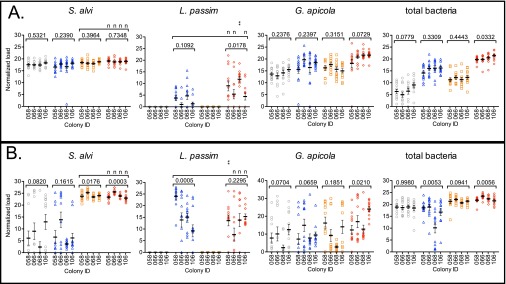
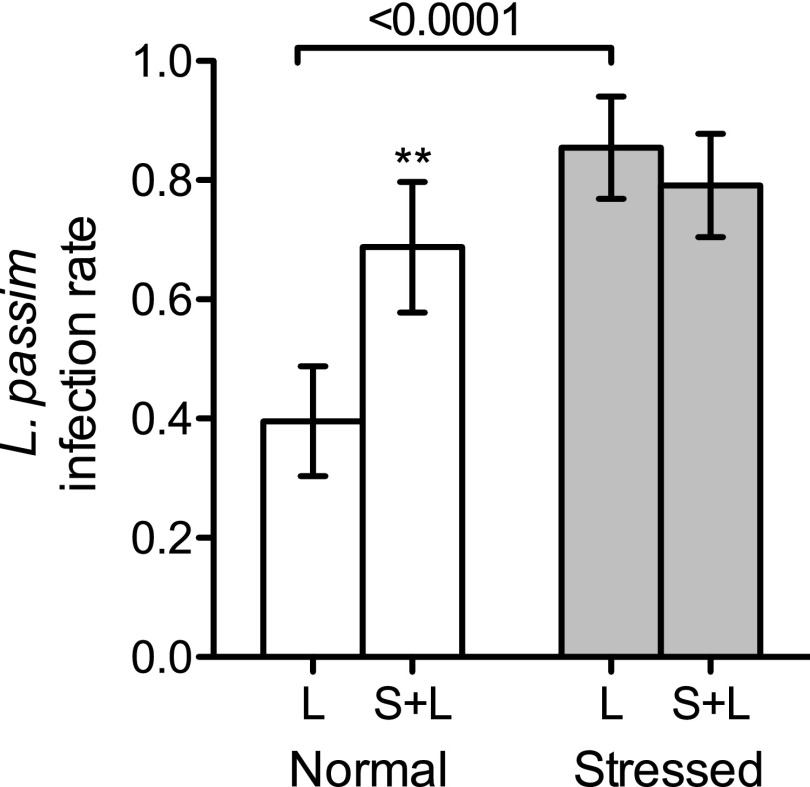
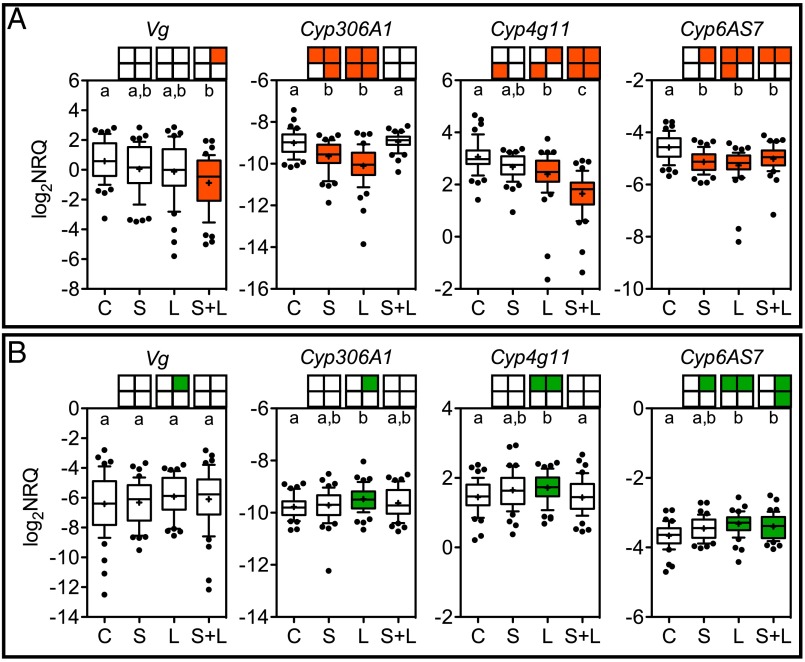
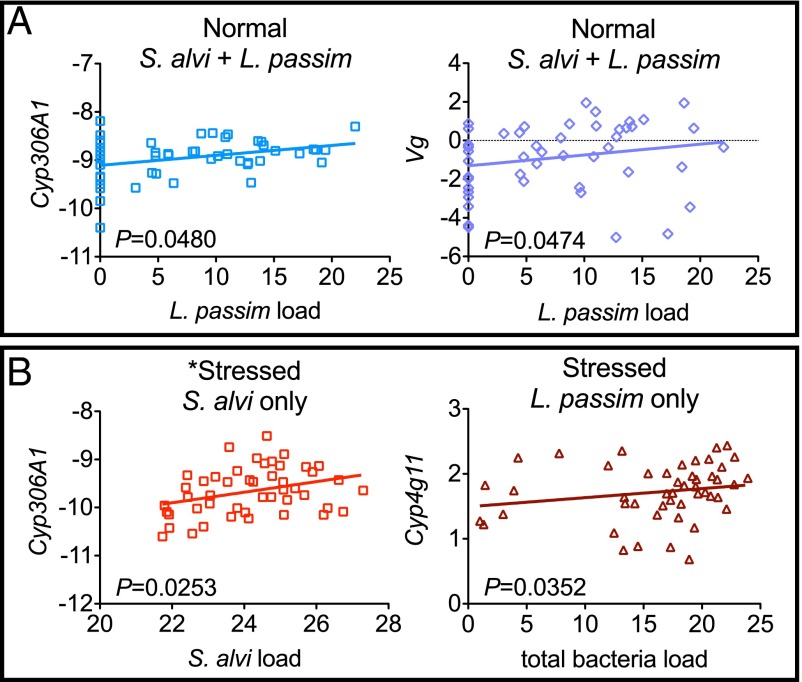
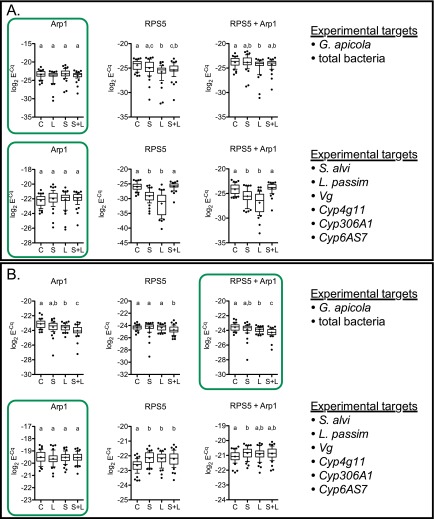
Microbial symbionts living within animal guts are largely composed of resident bacterial species, forming communities that often provide benefits to the host. Gut microbiomes of adult honey bees (Apis mellifera) include core residents such as the betaproteobacterium Snodgrassella alvi, alongside transient parasites such as the protozoan Lotmaria passim To test how these species affect microbiome composition and host physiology, we administered S alvi and/or L passim inocula to newly emerged worker bees from four genetic backgrounds (GH) and reared them in normal (within hives) or stressed (protein-deficient, asocial) conditions. Microbiota acquired by normal bees were abundant but quantitatively differed across treatments, indicating treatment-associated dysbiosis. Pretreatment with S. alvi made normal bees more susceptible to L. passim and altered developmental and detoxification gene expression. Stressed bees were more susceptible to L. passim and were depauperate in core microbiota, yet supplementation with S. alvi did not alter this susceptibility. Microbiomes were generally more variable by GH in stressed bees, which also showed opposing and comparatively reduced modulation of gene expression responses to treatments compared with normal bees. These data provide experimental support for a link between altered gut microbiota and increased parasite and pathogen prevalence, as observed from honey bee colony collapse disorder.
Keywords: cytochrome P450; dysbiosis; microbial interaction; parasitism; vitellogenin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Probiotic Treatment with a Gut Symbiont Leads to Parasite Susceptibility in Honey Bees.Trends Parasitol. 2016 Dec;32(12):914-916. doi: 10.1016/j.pt.2016.09.005. Epub 2016 Sep 30. Trends Parasitol. 2016. PMID: 27697444
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
