

miR-146b antagomir-treated human Tregs acquire increased GVHD inhibitory potency
- PMID: 27485827
- PMCID: PMC5016708
- DOI: 10.1182/blood-2016-05-714535
miR-146b antagomir-treated human Tregs acquire increased GVHD inhibitory potency
Abstract
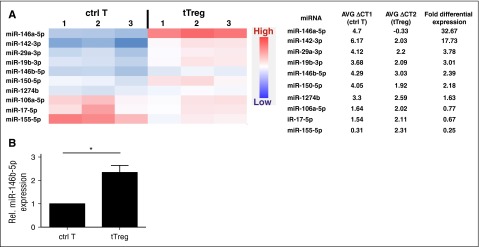
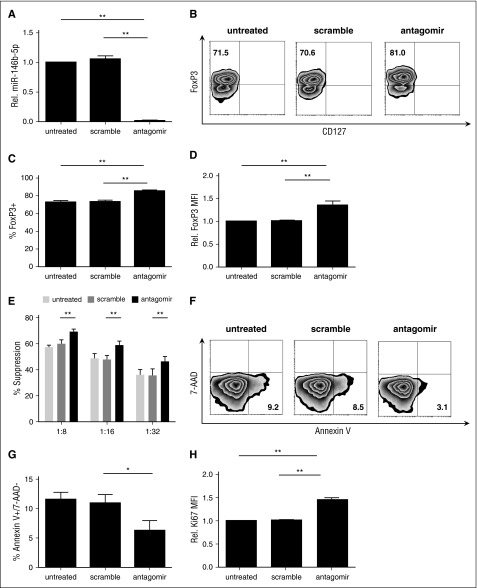
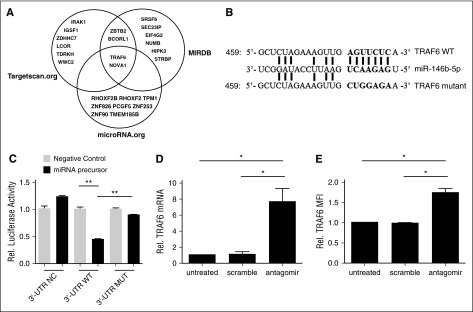
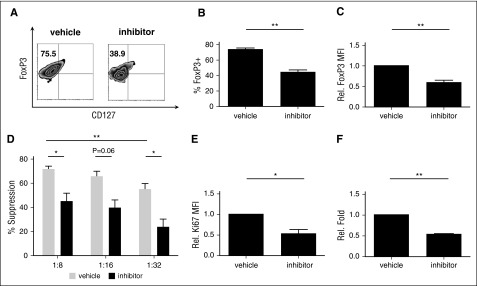
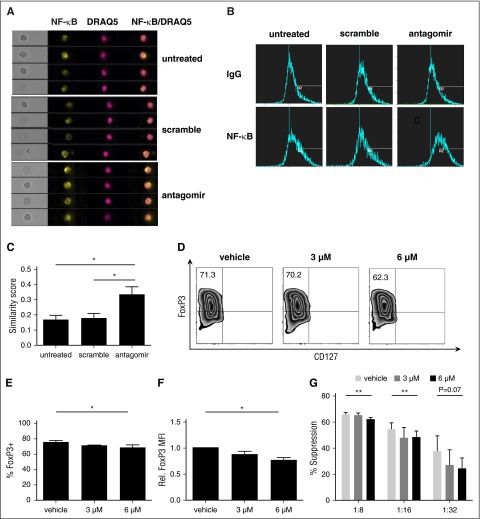
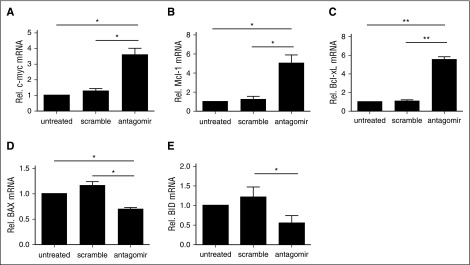
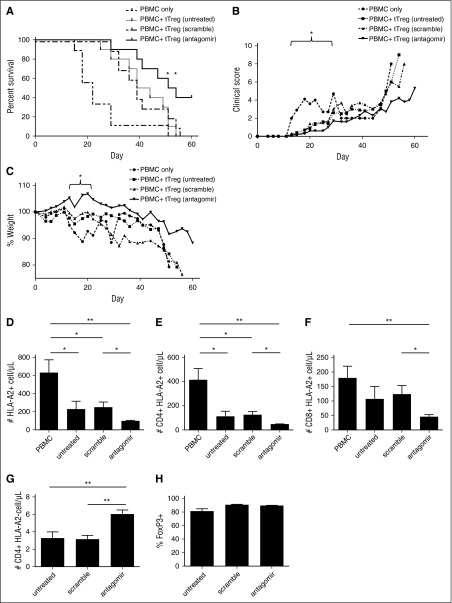
CD4(+)CD25(+)FoxP3(+) thymic-derived regulatory T cells (tTregs) are indispensable for maintaining immune system equilibrium. Adoptive transfer of tTregs is an effective means of suppressing graft-versus-host disease (GVHD) in murine models and in early human clinical trials. Tumor necrosis factor receptor-associated factor 6 (TRAF6), an ubiquitin-conjugating enzyme that mediates nuclear factor κB (NF-κB) activation, plays an essential role in modulating regulatory T cell survival and function. MicroRNAs (miRNAs) are noncoding RNAs, which mediate RNA silencing and posttranscriptional gene repression. By performing comprehensive TaqMan Low Density Array miRNA assays, we identified 10 miRNAs differentially regulated in human tTreg compared with control T cells. One candidate, miR-146b, is preferentially and highly expressed in human naive tTregs compared with naive CD4 T cells. miRNA prediction software revealed that TRAF6 was the one of the top 10 scored mRNAs involved tTreg function with the highest probability as a potential miR-146b target. Antagomir-mediated knockdown of miRNA-146b, but not another miRNA-146 family member (miRNA-146a), enhanced TRAF6 expression. TRAF6, in turn, increases NF-κB activation, which is essential for tTreg function as well as Foxp3 protein and antiapoptotic gene expression, and downregulates proapoptotic gene expression. miR-146b knockdown increased the nuclear localization and expression of genes regulated by NF-κB, which was associated with enhanced tTreg survival, proliferation, and suppressive function measured in vitro and in vivo. TRAF6 inhibition had the opposite effects. We conclude that an miR-146b-TRAF6-NF-κB-FoxP3 signaling pathway restrains regulatory T cell survival, proliferation, and suppressor function. In vitro exposure of human tTregs to miR-146b antagomirs can be exploited to improve the clinical efficacy of human adoptive tTreg transfer in a GVHD setting.
© 2016 by The American Society of Hematology.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
