

Ablation of STAT3 in the B Cell Compartment Restricts Gammaherpesvirus Latency In Vivo
- PMID: 27486189
- PMCID: PMC4981709
- DOI: 10.1128/mBio.00723-16
Ablation of STAT3 in the B Cell Compartment Restricts Gammaherpesvirus Latency In Vivo
Abstract
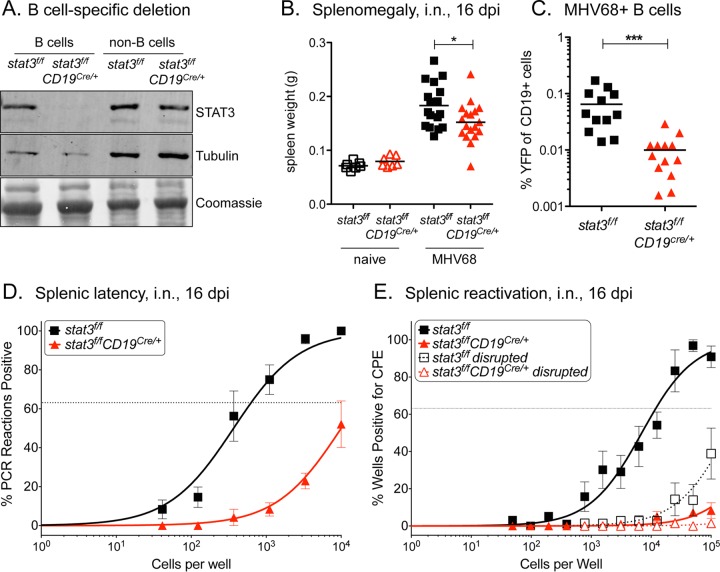
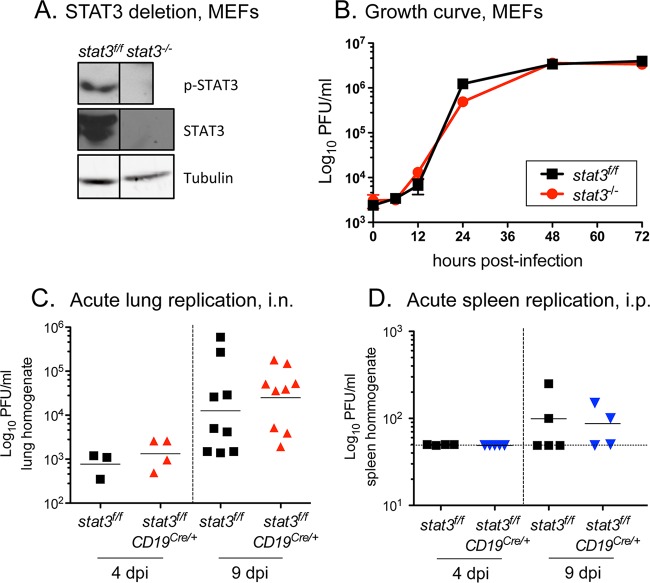
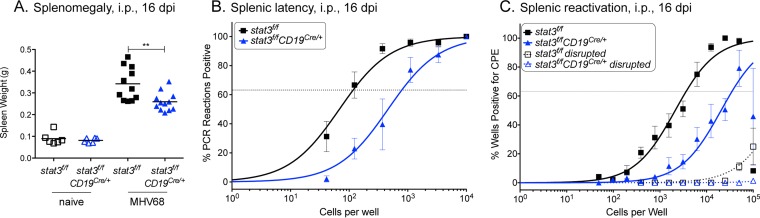
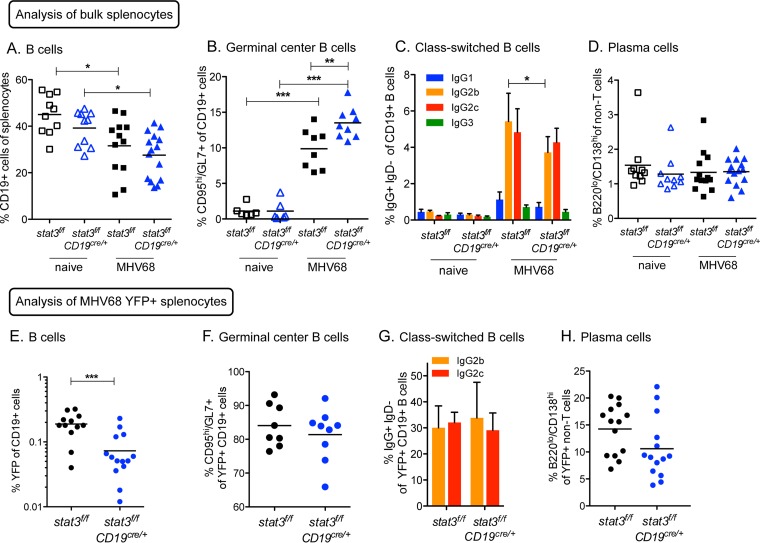
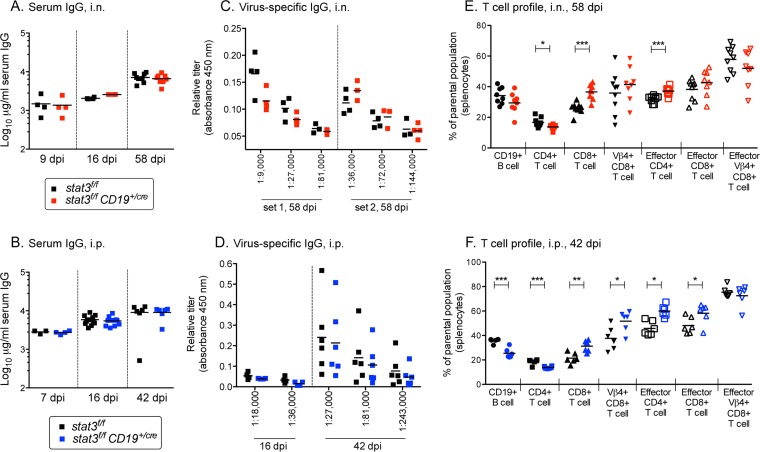
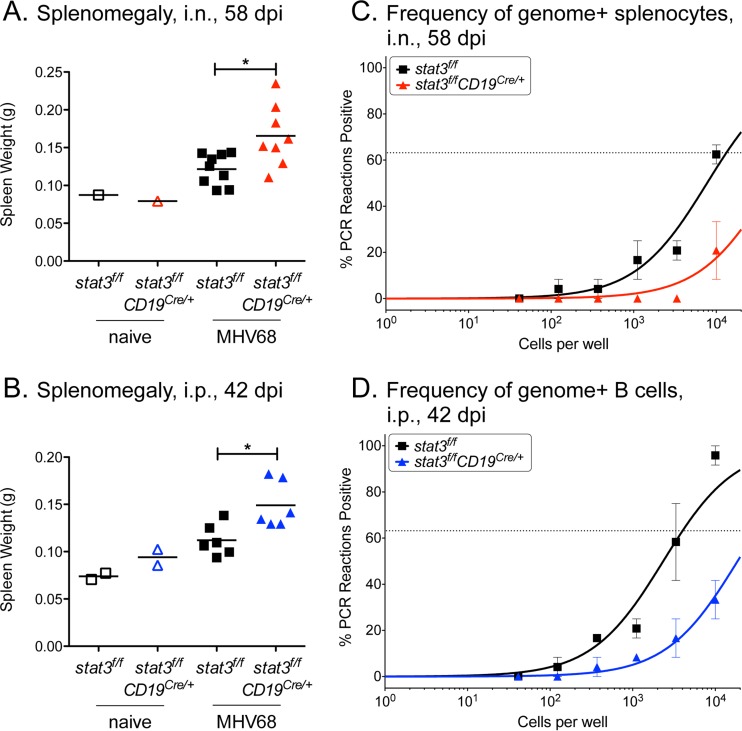
A challenging property of gammaherpesviruses is their ability to establish lifelong persistence. The establishment of latency in B cells is thought to involve active virus engagement of host signaling pathways. Pathogenic effects of these viruses during latency or following reactivation can be devastating to the host. Many cancers, including those associated with members of the gammaherpesvirus family, Kaposi's sarcoma-associated herpesvirus and Epstein-Barr virus, express elevated levels of active host signal transducer and activator of transcription-3 (STAT3). STAT3 is activated by tyrosine phosphorylation in response to many cytokines and can orchestrate effector responses that include proliferation, inflammation, metastasis, and developmental programming. However, the contribution of STAT3 to gammaherpesvirus pathogenesis remains to be completely understood. This is the first study to have identified STAT3 as a critical host determinant of the ability of gammaherpesvirus to establish long-term latency in an animal model of disease. Following an acute infection, murine gammaherpesvirus 68 (MHV68) established latency in resident B cells, but establishment of latency was dramatically reduced in animals with a B cell-specific STAT3 deletion. The lack of STAT3 in B cells did not impair germinal center responses for immunoglobulin (Ig) class switching in the spleen and did not reduce either total or virus-specific IgG titers. Although ablation of STAT3 in B cells did not have a global effect on these assays of B cell function, it had long-term consequences for the viral load of the host, since virus latency was reduced at 6 to 8 weeks postinfection. Our findings establish host STAT3 as a mediator of gammaherpesvirus persistence.
Importance: The insidious ability of gammaherpesviruses to establish latent infections can have detrimental consequences for the host. Identification of host factors that promote viral latency is essential for understanding latency mechanisms and for therapeutic interventions. We provide the first evidence that STAT3 expression is needed for murine gammaherpesvirus 68 to establish latency in primary B cells during an active immune response to infection. STAT3 deletion in B cells does not impair adaptive immune control of the virus, but loss of STAT3 in B cells has a long-lasting impact on viral persistence. These results indicate a potential therapeutic benefit of STAT3 inhibitors for combating gammaherpesvirus latency and, thereby, associated pathologies.
Copyright © 2016 Reddy et al.
Figures
Similar articles
-
Multifaceted roles for STAT3 in gammaherpesvirus latency revealed through in vivo B cell knockout models.mBio. 2024 Feb 14;15(2):e0299823. doi: 10.1128/mbio.02998-23. Epub 2024 Jan 3. mBio. 2024. PMID: 38170993 Free PMC article.
-
Deletion of Murine Gammaherpesvirus Gene M2 in Activation-Induced Cytidine Deaminase-Expressing B Cells Impairs Host Colonization and Viral Reactivation.J Virol. 2020 Dec 9;95(1):e01933-20. doi: 10.1128/JVI.01933-20. Print 2020 Dec 9. J Virol. 2020. PMID: 33028711 Free PMC article.
-
Murine gammaherpesvirus 68 reactivation from B cells requires IRF4 but not XBP-1.J Virol. 2014 Oct;88(19):11600-10. doi: 10.1128/JVI.01876-14. Epub 2014 Jul 30. J Virol. 2014. PMID: 25078688 Free PMC article.
-
Murine Gammaherpesvirus 68: A Small Animal Model for Gammaherpesvirus-Associated Diseases.Adv Exp Med Biol. 2017;1018:225-236. doi: 10.1007/978-981-10-5765-6_14. Adv Exp Med Biol. 2017. PMID: 29052141 Review.
-
Interplay of Murine Gammaherpesvirus 68 with NF-kappaB Signaling of the Host.Front Microbiol. 2016 Aug 17;7:1202. doi: 10.3389/fmicb.2016.01202. eCollection 2016. Front Microbiol. 2016. PMID: 27582728 Free PMC article. Review.
Cited by
-
The regulatory role of protein phosphorylation in human gammaherpesvirus associated cancers.Virol Sin. 2017 Oct;32(5):357-368. doi: 10.1007/s12250-017-4081-9. Epub 2017 Oct 30. Virol Sin. 2017. PMID: 29116588 Free PMC article. Review.
-
B cell-intrinsic STAT3-mediated support of latency and interferon suppression during murine gammaherpesvirus 68 infection revealed through an in vivo competition model.bioRxiv [Preprint]. 2023 Mar 22:2023.03.22.533727. doi: 10.1101/2023.03.22.533727. bioRxiv. 2023. PMID: 36993230 Free PMC article. Preprint.
-
Interferon Regulatory Factor 3 Supports the Establishment of Chronic Gammaherpesvirus Infection in a Route- and Dose-Dependent Manner.J Virol. 2021 Apr 12;95(9):e02208-20. doi: 10.1128/JVI.02208-20. Print 2021 Apr 12. J Virol. 2021. PMID: 33597211 Free PMC article.
-
Conquering the Host: Determinants of Pathogenesis Learned from Murine Gammaherpesvirus 68.Annu Rev Virol. 2021 Sep 29;8(1):349-371. doi: 10.1146/annurev-virology-011921-082615. Annu Rev Virol. 2021. PMID: 34586873 Free PMC article.
-
Germinal center cytokine driven epigenetic control of Epstein-Barr virus latency gene expression.PLoS Pathog. 2024 Apr 29;20(4):e1011939. doi: 10.1371/journal.ppat.1011939. eCollection 2024 Apr. PLoS Pathog. 2024. PMID: 38683861 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
