

Dishevelled is a NEK2 kinase substrate controlling dynamics of centrosomal linker proteins
- PMID: 27486244
- PMCID: PMC4995965
- DOI: 10.1073/pnas.1608783113
Dishevelled is a NEK2 kinase substrate controlling dynamics of centrosomal linker proteins
Abstract
Dishevelled (DVL) is a key scaffolding protein and a branching point in Wnt signaling pathways. Here, we present conclusive evidence that DVL regulates the centrosomal cycle. We demonstrate that DVL dishevelled and axin (DIX) domain, but not DIX domain-mediated multimerization, is essential for DVL's centrosomal localization. DVL accumulates during the cell cycle and associates with NIMA-related kinase 2 (NEK2), which is able to phosphorylate DVL at a multitude of residues, as detected by a set of novel phospho-specific antibodies. This creates interfaces for efficient binding to CDK5 regulatory subunit-associated protein 2 (CDK5RAP2) and centrosomal Nek2-associated protein 1 (C-NAP1), two proteins of the centrosomal linker. Displacement of DVL from the centrosome and its release into the cytoplasm on NEK2 phosphorylation is coupled to the removal of linker proteins, an event necessary for centrosomal separation and proper formation of the mitotic spindle. Lack of DVL prevents NEK2-controlled dissolution of loose centrosomal linker and subsequent centrosomal separation. Increased DVL levels, in contrast, sequester centrosomal NEK2 and mimic monopolar spindle defects induced by a dominant negative version of this kinase. Our study thus uncovers molecular crosstalk between centrosome and Wnt signaling.
Keywords: Dishevelled; NEK2; Wnt signaling; centrosome; linker proteins.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
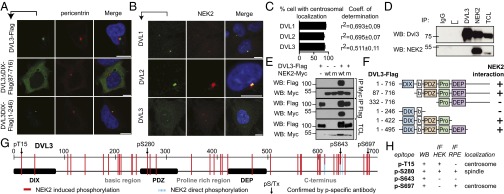
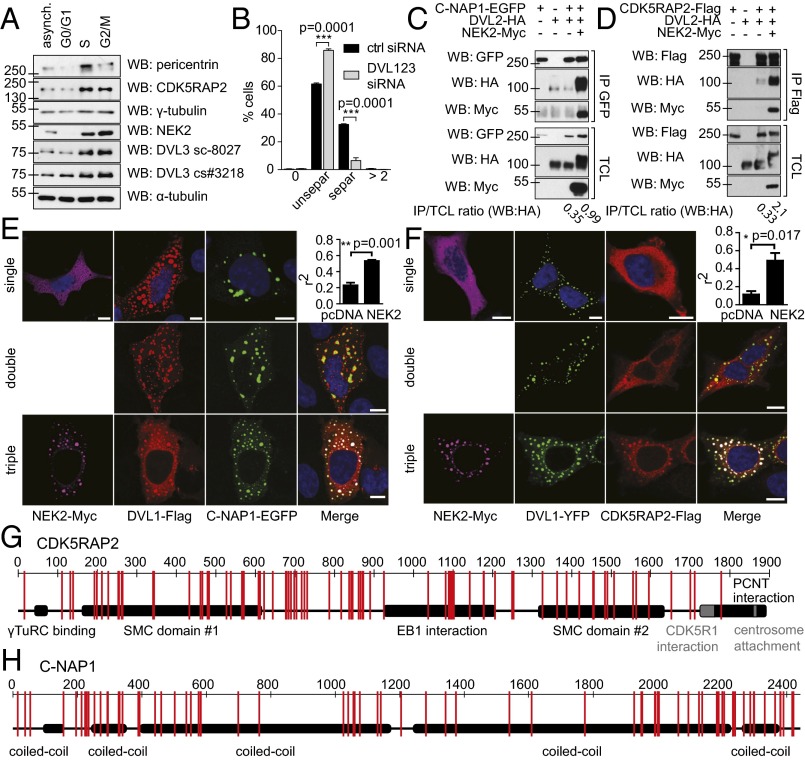
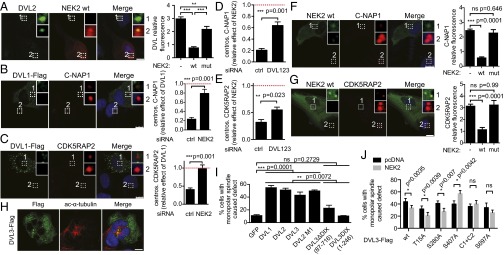
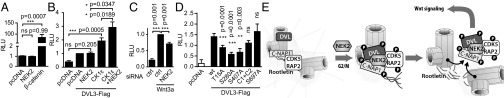
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
