

The stringent response regulates adaptation to darkness in the cyanobacterium Synechococcus elongatus
- PMID: 27486247
- PMCID: PMC4995992
- DOI: 10.1073/pnas.1524915113
The stringent response regulates adaptation to darkness in the cyanobacterium Synechococcus elongatus
Abstract
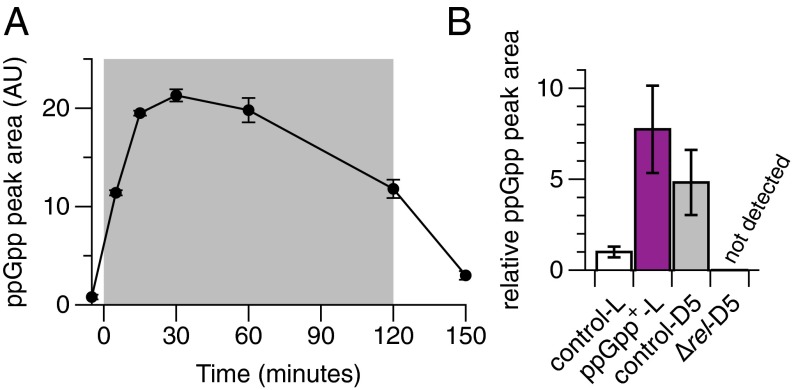
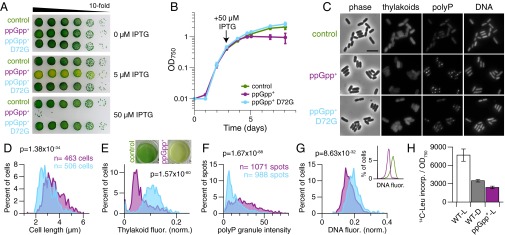
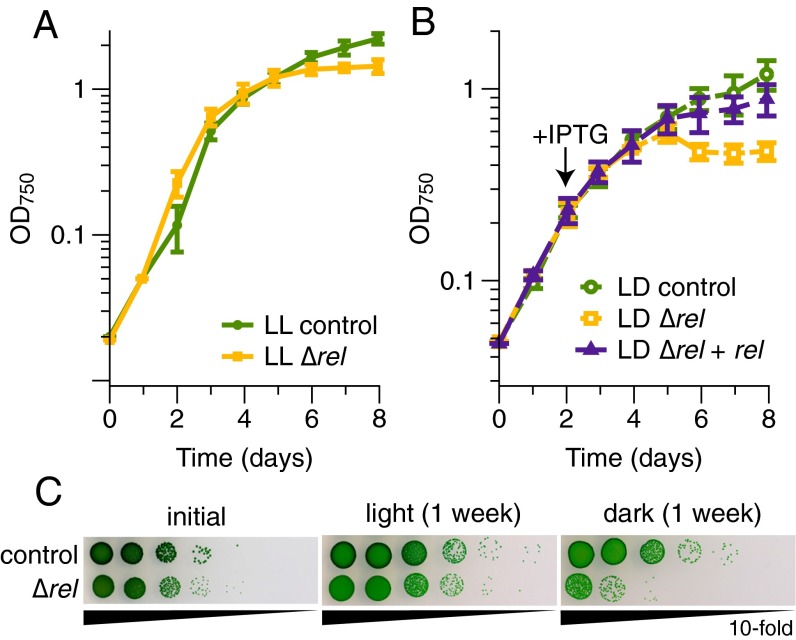
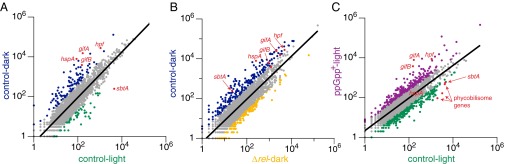
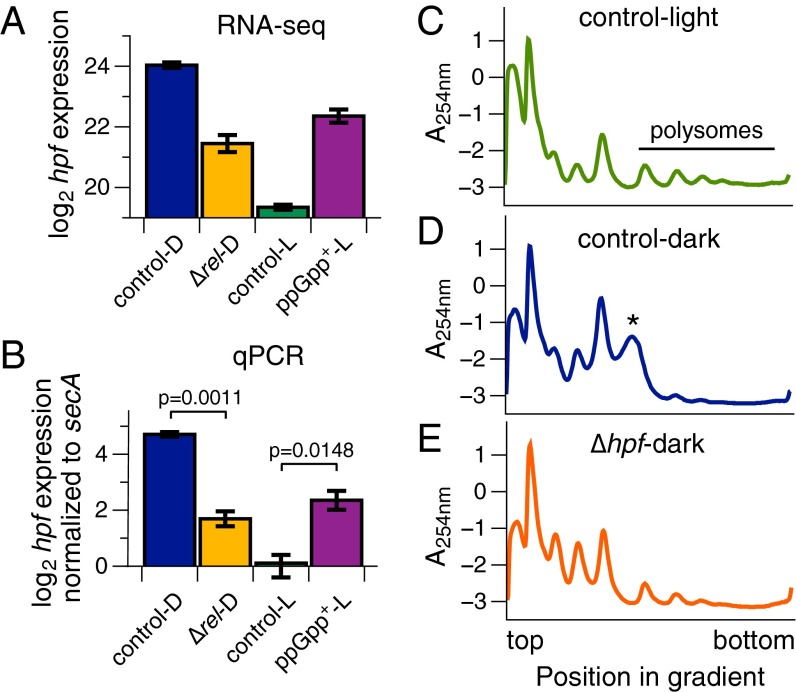
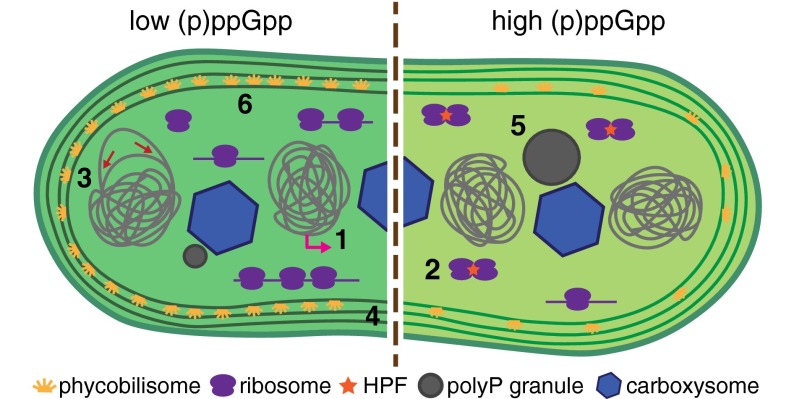
The cyanobacterium Synechococcus elongatus relies upon photosynthesis to drive metabolism and growth. During darkness, Synechococcus stops growing, derives energy from its glycogen stores, and greatly decreases rates of macromolecular synthesis via unknown mechanisms. Here, we show that the stringent response, a stress response pathway whose genes are conserved across bacteria and plant plastids, contributes to this dark adaptation. Levels of the stringent response alarmone guanosine 3'-diphosphate 5'-diphosphate (ppGpp) rise after a shift from light to dark, indicating that darkness triggers the same response in cyanobacteria as starvation in heterotrophic bacteria. High levels of ppGpp are sufficient to stop growth and dramatically alter many aspects of cellular physiology, including levels of photosynthetic pigments and polyphosphate, DNA content, and the rate of translation. Cells unable to synthesize ppGpp display pronounced growth defects after exposure to darkness. The stringent response regulates expression of a number of genes in Synechococcus, including ribosomal hibernation promoting factor (hpf), which causes ribosomes to dimerize in the dark and may contribute to decreased translation. Although the metabolism of Synechococcus differentiates it from other model bacterial systems, the logic of the stringent response remains remarkably conserved, while at the same time having adapted to the unique stresses of the photosynthetic lifestyle.
Keywords: (p)ppGpp; Synechococcus; cyanobacteria; hibernation promoting factor; stringent response.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
ppGpp Controls Global Gene Expression in Light and in Darkness in S. elongatus.Cell Rep. 2017 Dec 12;21(11):3155-3165. doi: 10.1016/j.celrep.2017.11.067. Cell Rep. 2017. PMID: 29241543
-
Effects of light deprivation on RNA synthesis, accumulation of guanosine 3'(2')-diphosphate 5'-diphosphate, and protein synthesis in heat-shocked Synechococcus sp. strain PCC 6301, a cyanobacterium.J Bacteriol. 1987 Feb;169(2):632-9. doi: 10.1128/jb.169.2.632-639.1987. J Bacteriol. 1987. PMID: 2433265 Free PMC article.
-
Physiological analysis of the stringent response elicited in an extreme thermophilic bacterium, Thermus thermophilus.J Bacteriol. 2006 Oct;188(20):7111-22. doi: 10.1128/JB.00574-06. J Bacteriol. 2006. PMID: 17015650 Free PMC article.
-
The synthesis and function of the alarmone (p)ppGpp in firmicutes.Int J Med Microbiol. 2010 Feb;300(2-3):142-7. doi: 10.1016/j.ijmm.2009.08.017. Epub 2009 Sep 24. Int J Med Microbiol. 2010. PMID: 19783475 Review.
-
Direct binding targets of the stringent response alarmone (p)ppGpp.Mol Microbiol. 2012 Sep;85(6):1029-43. doi: 10.1111/j.1365-2958.2012.08177.x. Epub 2012 Aug 2. Mol Microbiol. 2012. PMID: 22812515 Review.
Cited by
-
Allosteric regulation of glycogen breakdown by the second messenger cyclic di-GMP.Nat Commun. 2022 Oct 3;13(1):5834. doi: 10.1038/s41467-022-33537-w. Nat Commun. 2022. PMID: 36192422 Free PMC article.
-
Transcriptional Responses to ppGpp and DksA.Annu Rev Microbiol. 2018 Sep 8;72:163-184. doi: 10.1146/annurev-micro-090817-062444. Annu Rev Microbiol. 2018. PMID: 30200857 Free PMC article. Review.
-
ATP Is a Major Determinant of Phototrophic Bacterial Longevity in Growth Arrest.mBio. 2023 Apr 25;14(2):e0360922. doi: 10.1128/mbio.03609-22. Epub 2023 Feb 14. mBio. 2023. PMID: 36786592 Free PMC article.
-
Disassembly of the Staphylococcus aureus hibernating 100S ribosome by an evolutionarily conserved GTPase.Proc Natl Acad Sci U S A. 2017 Sep 26;114(39):E8165-E8173. doi: 10.1073/pnas.1709588114. Epub 2017 Sep 11. Proc Natl Acad Sci U S A. 2017. PMID: 28894000 Free PMC article.
-
Hibernation factors directly block ribonucleases from entering the ribosome in response to starvation.Nucleic Acids Res. 2021 Feb 26;49(4):2226-2239. doi: 10.1093/nar/gkab017. Nucleic Acids Res. 2021. PMID: 33503254 Free PMC article.
References
-
- Külheim C, Agren J, Jansson S. Rapid regulation of light harvesting and plant fitness in the field. Science. 2002;297(5578):91–93. - PubMed
-
- Bailey S, Grossman A. Photoprotection in cyanobacteria: Regulation of light harvesting. Photochem Photobiol. 2008;84(6):1410–1420. - PubMed
-
- Doolittle WF. The cyanobacterial genome, its expression, and the control of that expression. Adv Microb Physiol. 1979;20:1–102. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
