

KLF10 Mediated Epigenetic Dysregulation of Epithelial CD40/CD154 Promotes Endometriosis
- PMID: 27488034
- PMCID: PMC5333936
- DOI: 10.1095/biolreprod.116.140764
KLF10 Mediated Epigenetic Dysregulation of Epithelial CD40/CD154 Promotes Endometriosis
Abstract
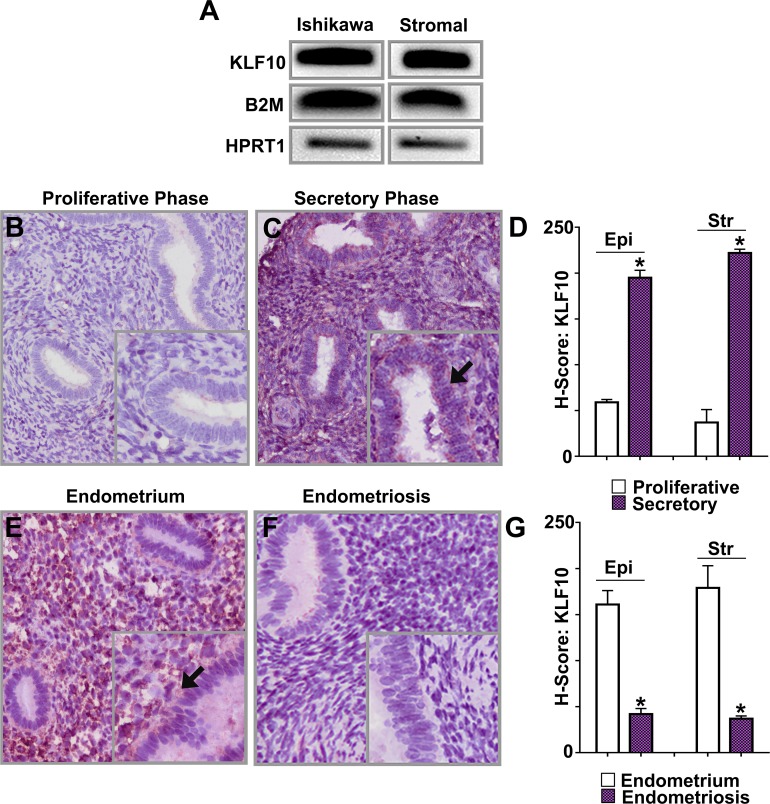
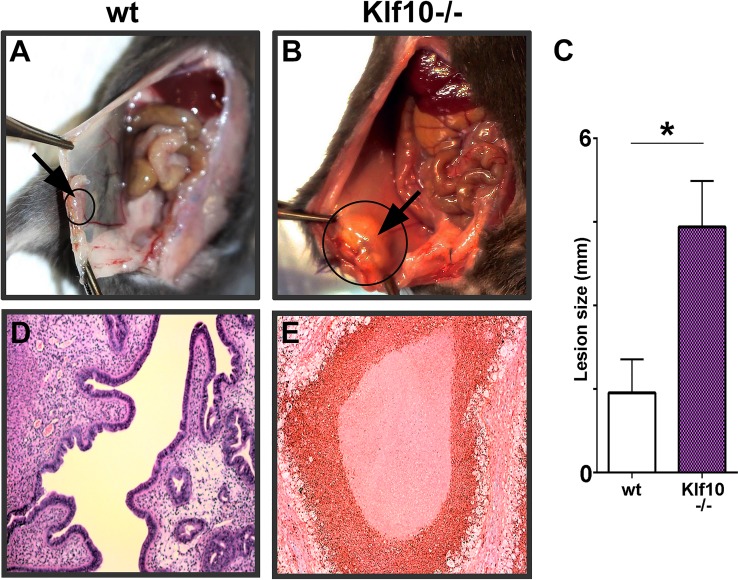
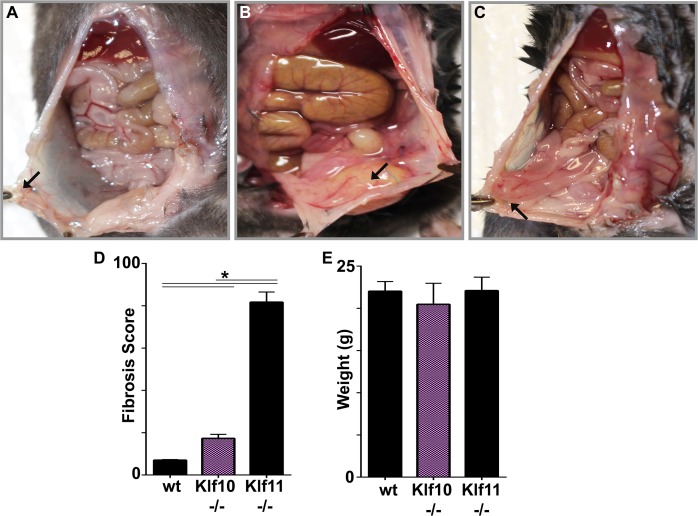
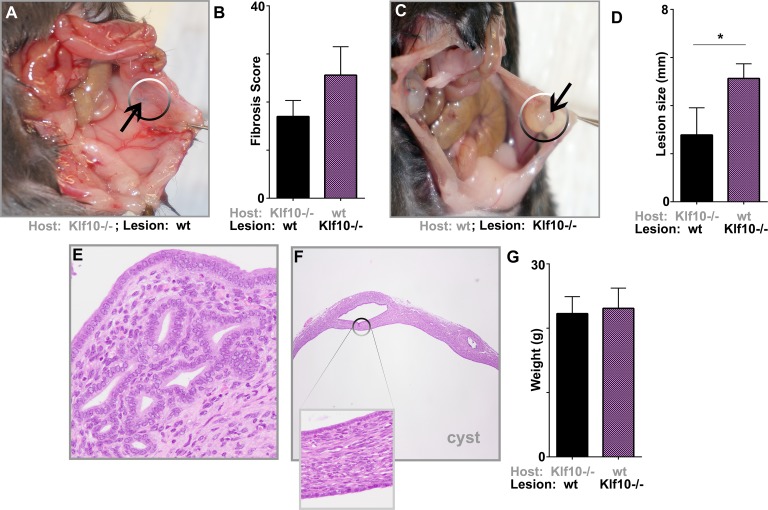
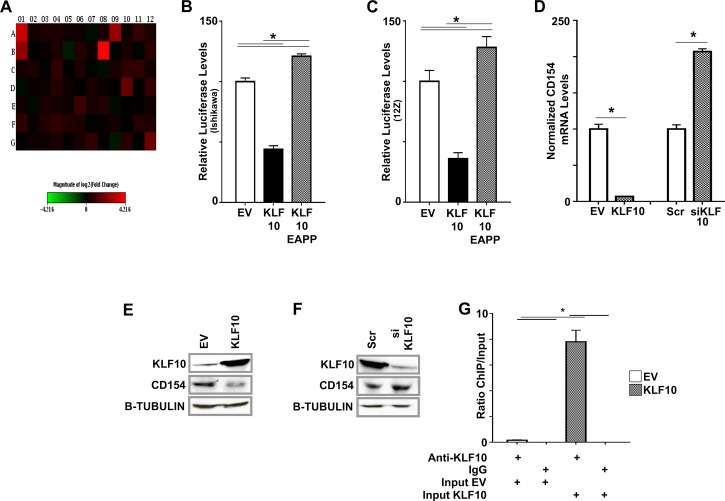
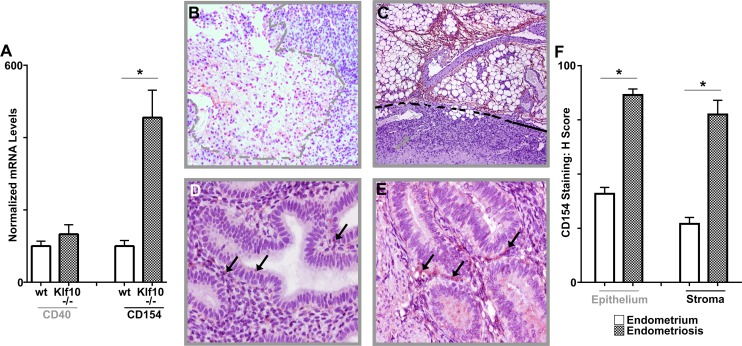
Endometriosis is a highly prevalent, chronic, heterogeneous, fibro-inflammatory disease that remains recalcitrant to conventional therapy. We previously showed that loss of KLF11, a transcription factor implicated in uterine disease, results in progression of endometriosis. Despite extensive homology, co-expression, and human disease association, loss of the paralog Klf10 causes a unique inflammatory, cystic endometriosis phenotype in contrast to fibrotic progression seen with loss of Klf11. We identify here for the first time a novel role for KLF10 in endometriosis. In an animal endometriosis model, unlike wild-type controls, Klf10(-/-) animals developed cystic lesions with massive immune infiltrate and minimal peri-lesional fibrosis. The Klf10(-/-) disease progression phenotype also contrasted with prolific fibrosis and minimal immune cell infiltration seen in Klf11(-/-) animals. We further found that lesion genotype rather than that of the host determined each unique disease progression phenotype. Mechanistically, KLF10 regulated CD40/CD154-mediated immune pathways. Both inflammatory as well as fibrotic phenotypes are the commonest clinical manifestations in chronic fibro-inflammatory diseases such as endometriosis. The complementary, paralogous Klf10 and Klf11 models therefore offer novel insights into the mechanisms of inflammation and fibrosis in a disease-relevant context. Our data suggests that divergence in underlying gene dysregulation critically determines disease-phenotype predominance rather than the conventional paradigm of inflammation being precedent to fibrotic scarring. Heterogeneity in clinical progression and treatment response are thus likely from disparate gene regulation profiles. Characterization of disease phenotype-associated gene dysregulation offers novel approaches for developing targeted, individualized therapy for recurrent and recalcitrant chronic disease.
Keywords: CD154; CD40; CD40 ligand; KLF10; endometriosis; epigenetics; inflammation.
© 2016 by the Society for the Study of Reproduction, Inc.
Figures
References
-
- Giudice LC, Kao LC. Endometriosis. Lancet. 2004;364:1789–1799. - PubMed
-
- Bonnefond A, Lomberk G, Buttar N, Busiah K, Vaillant E, Lobbens S, Yengo L, Dechaume A, Mignot B, Simon A, Scharfmann R, Neve B, et al. Disruption of a novel Kruppel-like transcription factor p300-regulated pathway for insulin biosynthesis revealed by studies of the c.-331 INS mutation found in neonatal diabetes mellitus. J Biol Chem. 2011;286:28414–28424. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
