

Dissecting relative contributions of cis- and trans-determinants to nucleosome distribution by comparing Tetrahymena macronuclear and micronuclear chromatin
- PMID: 27488188
- PMCID: PMC5137421
- DOI: 10.1093/nar/gkw684
Dissecting relative contributions of cis- and trans-determinants to nucleosome distribution by comparing Tetrahymena macronuclear and micronuclear chromatin
Abstract
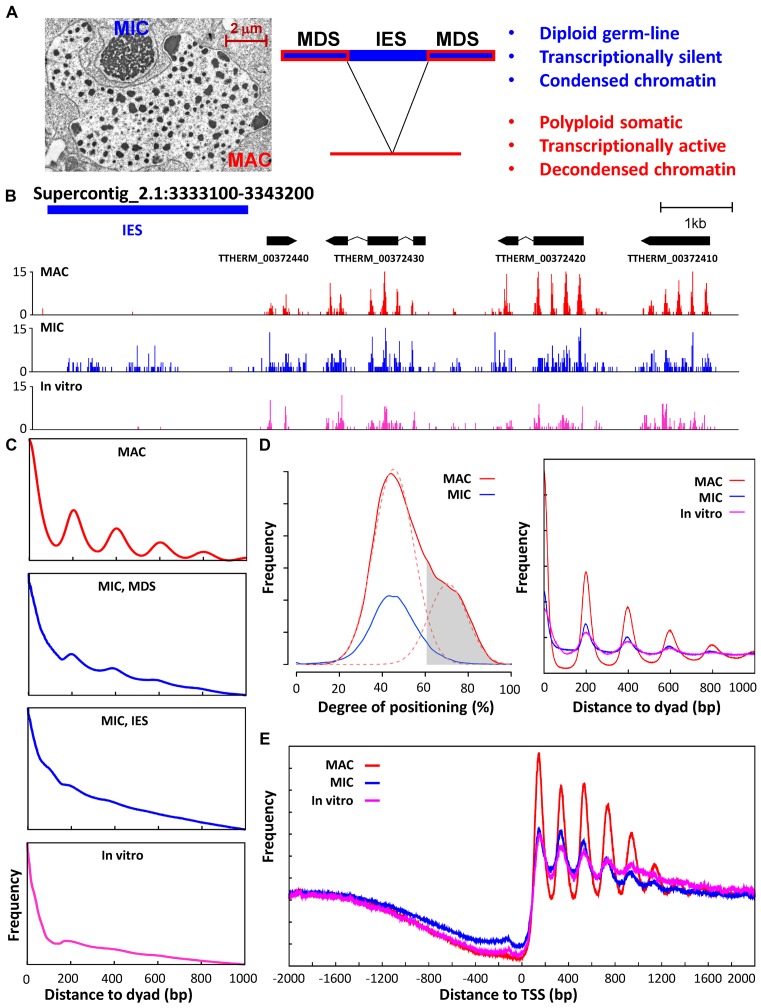
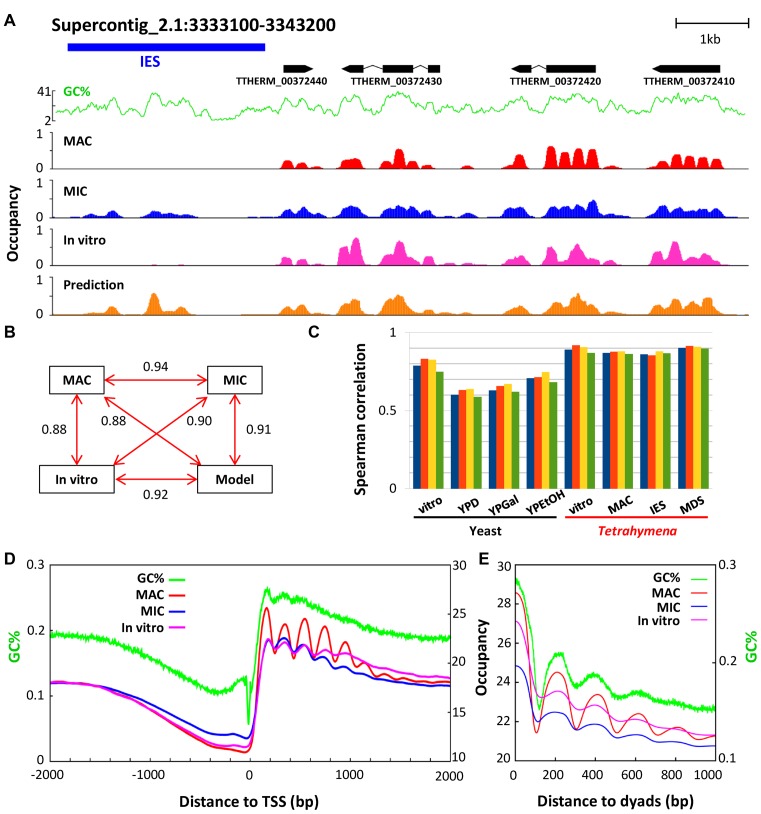
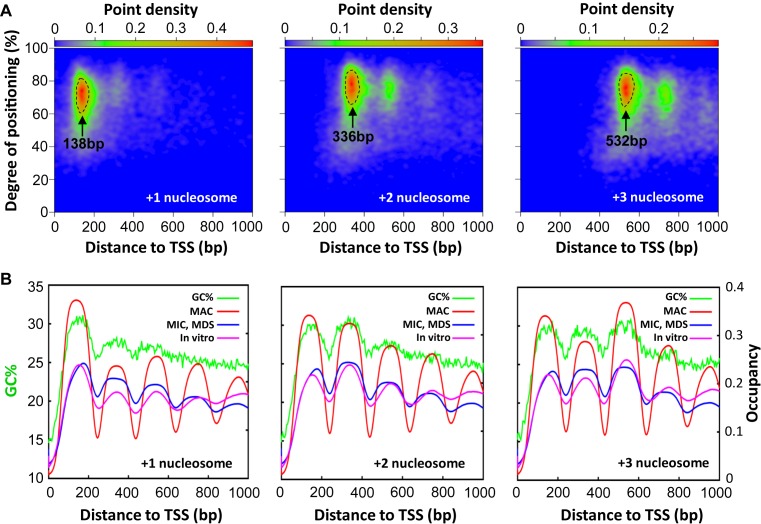
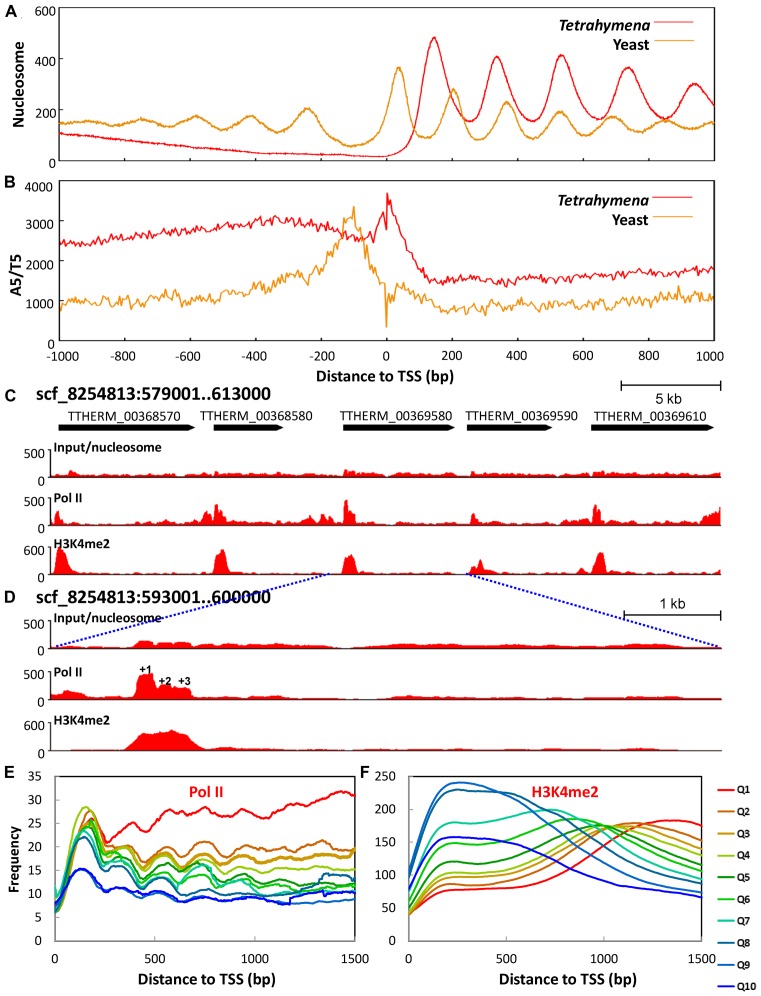
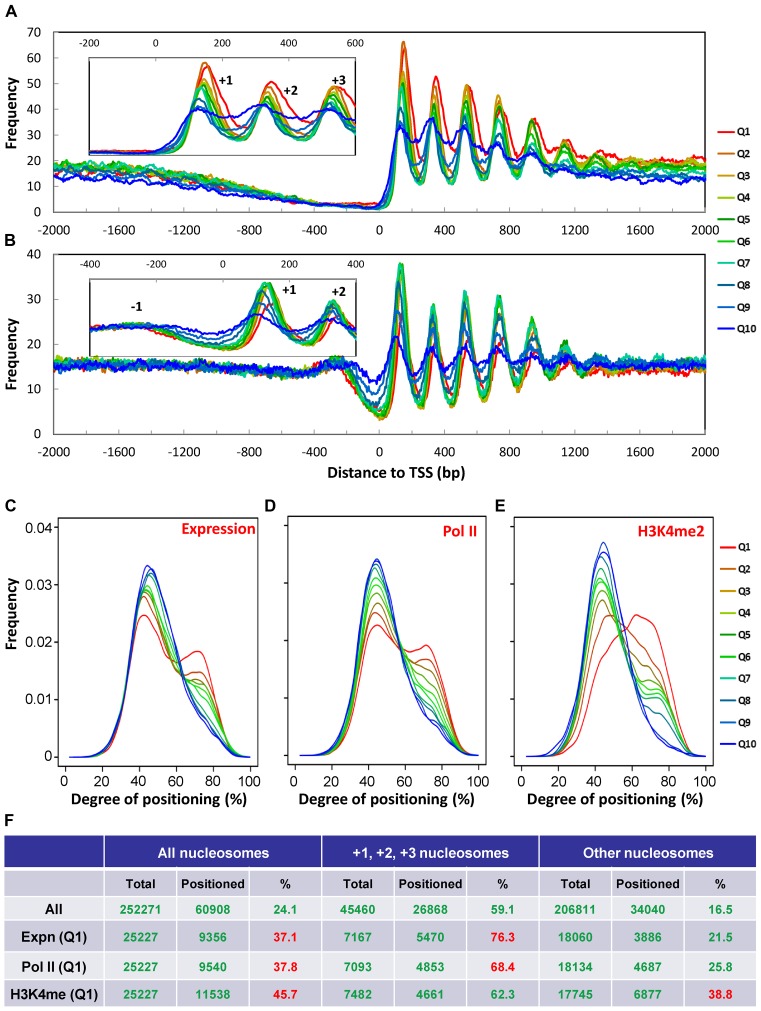
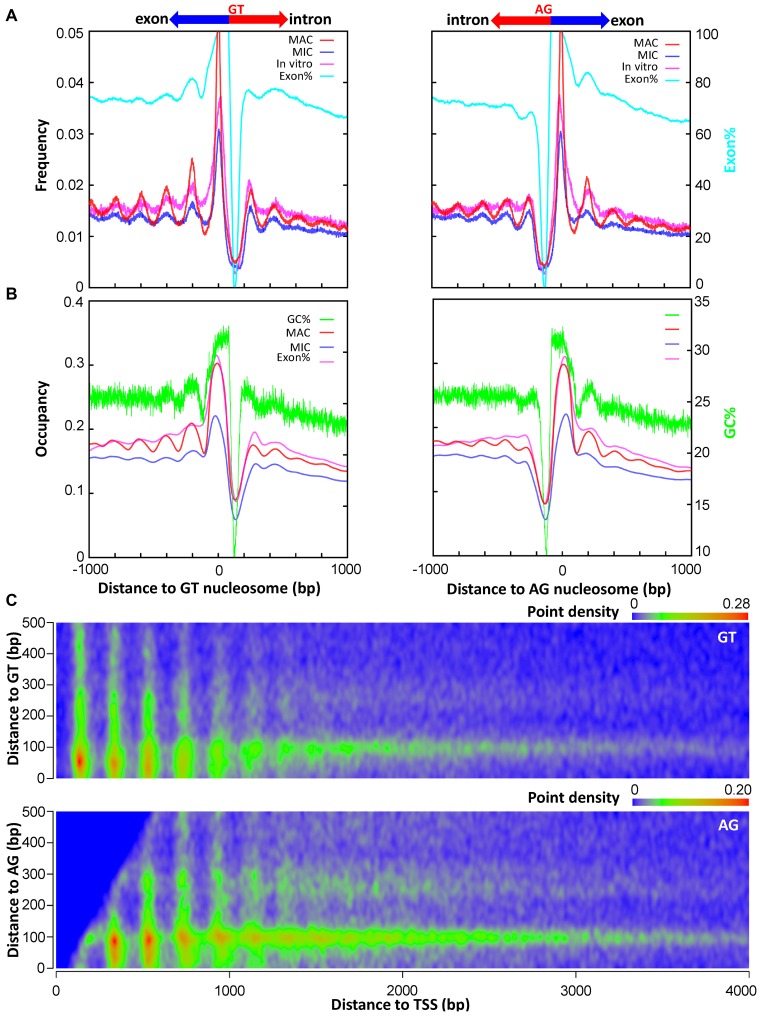
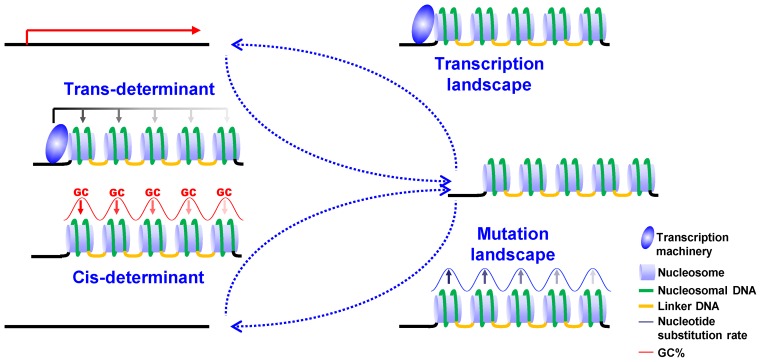
The ciliate protozoan Tetrahymena thermophila contains two types of structurally and functionally differentiated nuclei: the transcriptionally active somatic macronucleus (MAC) and the transcriptionally silent germ-line micronucleus (MIC). Here, we demonstrate that MAC features well-positioned nucleosomes downstream of transcription start sites and flanking splice sites. Transcription-associated trans-determinants promote nucleosome positioning in MAC. By contrast, nucleosomes in MIC are dramatically delocalized. Nucleosome occupancy in MAC and MIC are nonetheless highly correlated with each other, as well as with in vitro reconstitution and predictions based upon DNA sequence features, revealing unexpectedly strong contributions from cis-determinants. In particular, well-positioned nucleosomes are often matched with GC content oscillations. As many nucleosomes are coordinately accommodated by both cis- and trans-determinants, we propose that their distribution is shaped by the impact of these nucleosomes on the mutational and transcriptional landscape, and driven by evolutionary selection.
© The Author(s) 2016. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Kornberg R.D., Lorch Y. Twenty-five years of the nucleosome, fundamental particle of the eukaryote chromosome. Cell. 1999;98:285–294. - PubMed
-
- Hughes A.L., Rando O.J. Mechanisms underlying nucleosome positioning in vivo. Annu. Rev. Biophysi. 2014;43:41–63. - PubMed
-
- Albert I., Mavrich T.N., Tomsho L.P., Qi J., Zanton S.J., Schuster S.C., Pugh B.F. Translational and rotational settings of H2A.Z nucleosomes across the Saccharomyces cerevisiae genome. Nature. 2007;446:572–576. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
