

Sensory neuropathy in progressive motor neuronopathy (pmn) mice is associated with defects in microtubule polymerization and axonal transport
- PMID: 27488538
- PMCID: PMC8029086
- DOI: 10.1111/bpa.12422
Sensory neuropathy in progressive motor neuronopathy (pmn) mice is associated with defects in microtubule polymerization and axonal transport
Abstract
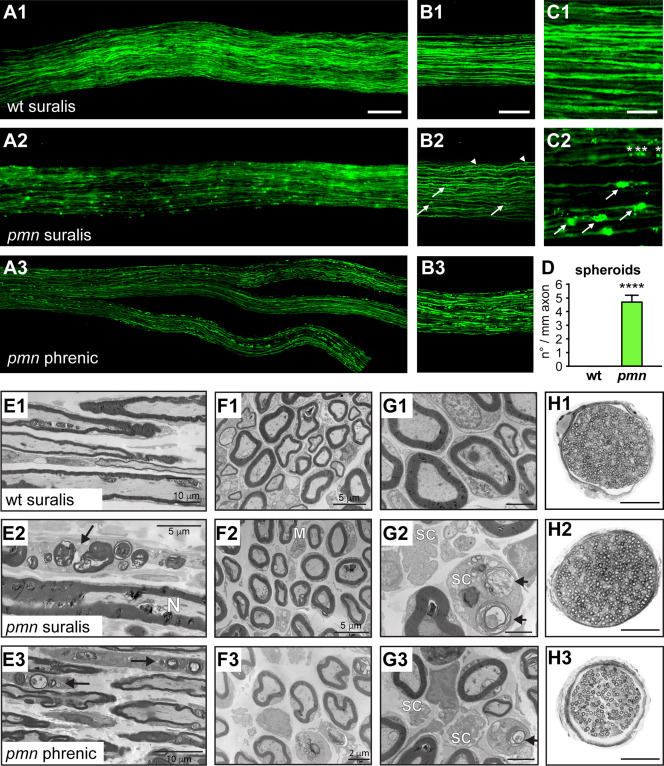
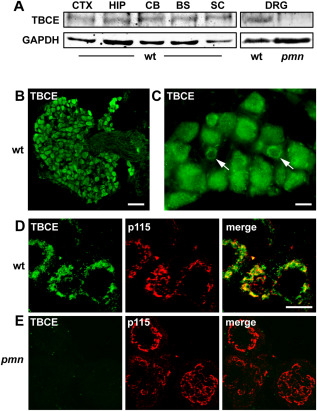
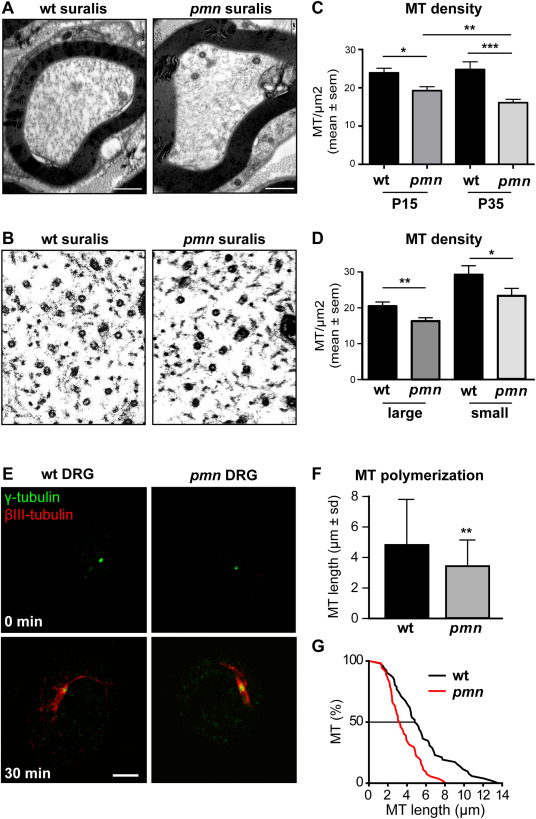
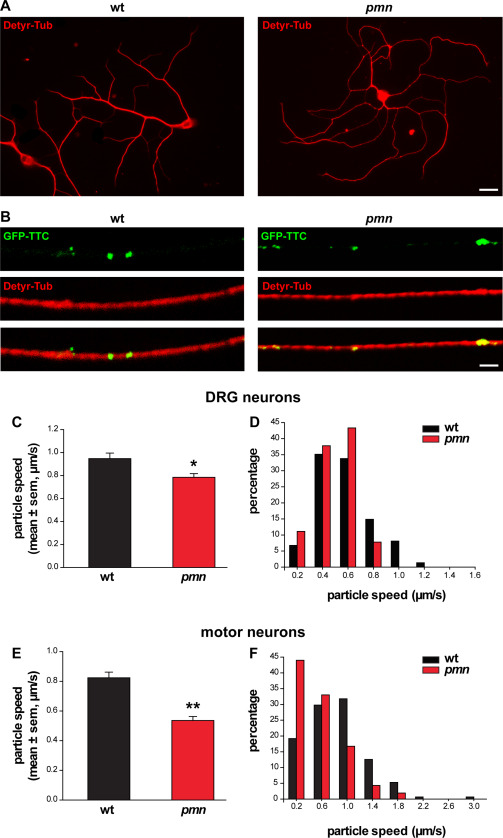
Motor neuron diseases such as amyotrophic lateral sclerosis (ALS) are now recognized as multi-system disorders also involving various non-motor neuronal cell types. The precise extent and mechanistic basis of non-motor neuron damage in human ALS and ALS animal models remain however unclear. To address this, we here studied progressive motor neuronopathy (pmn) mice carrying a missense loss-of-function mutation in tubulin binding cofactor E (TBCE). These mice manifest a particularly aggressive form of motor axon dying back and display a microtubule loss, similar to that induced by human ALS-linked TUBA4A mutations. Using whole nerve confocal imaging of pmn × thy1.2-YFP16 fluorescent reporter mice and electron microscopy, we demonstrate axonal discontinuities, bead-like spheroids and ovoids in pmn suralis nerves indicating prominent sensory neuropathy. The axonal alterations qualitatively resemble those in phrenic motor nerves but do not culminate in the loss of myelinated fibers. We further show that the pmn mutation decreases the level of TBCE, impedes microtubule polymerization in dorsal root ganglion (DRG) neurons and causes progressive loss of microtubules in large and small caliber suralis axons. Live imaging of axonal transport using GFP-tagged tetanus toxin C-fragment (GFP-TTC) demonstrates defects in microtubule-based transport in pmn DRG neurons, providing a potential explanation for the axonal alterations in sensory nerves. This study unravels sensory neuropathy as a pathological feature of mouse pmn, and discusses the potential contribution of cytoskeletal defects to sensory neuropathy in human motor neuron disease.
Keywords: axon degeneration; microtubule; motor neuron disease; sensory neuropathy; spheroid.
© 2016 International Society of Neuropathology.
Figures
Similar articles
-
Progressive motor neuronopathy: a critical role of the tubulin chaperone TBCE in axonal tubulin routing from the Golgi apparatus.J Neurosci. 2007 Aug 15;27(33):8779-89. doi: 10.1523/JNEUROSCI.1599-07.2007. J Neurosci. 2007. PMID: 17699660 Free PMC article.
-
Neurofilament depletion improves microtubule dynamics via modulation of Stat3/stathmin signaling.Acta Neuropathol. 2016 Jul;132(1):93-110. doi: 10.1007/s00401-016-1564-y. Epub 2016 Mar 28. Acta Neuropathol. 2016. PMID: 27021905 Free PMC article.
-
Missense mutation in the tubulin-specific chaperone E (Tbce) gene in the mouse mutant progressive motor neuronopathy, a model of human motoneuron disease.J Cell Biol. 2002 Nov 25;159(4):563-9. doi: 10.1083/jcb.200208001. J Cell Biol. 2002. PMID: 12446740 Free PMC article.
-
Golgi Fragmentation in ALS Motor Neurons. New Mechanisms Targeting Microtubules, Tethers, and Transport Vesicles.Front Neurosci. 2015 Dec 8;9:448. doi: 10.3389/fnins.2015.00448. eCollection 2015. Front Neurosci. 2015. PMID: 26696811 Free PMC article. Review.
-
Disruption of axonal transport in motor neuron diseases.Int J Mol Sci. 2012;13(1):1225-1238. doi: 10.3390/ijms13011225. Epub 2012 Jan 23. Int J Mol Sci. 2012. PMID: 22312314 Free PMC article. Review.
Cited by
-
Pegylated Insulin-Like Growth Factor 1 attenuates Hair Cell Loss and promotes Presynaptic Maintenance of Medial Olivocochlear Cholinergic Fibers in the Cochlea of the Progressive Motor Neuropathy Mouse.Front Neurol. 2022 Jun 3;13:885026. doi: 10.3389/fneur.2022.885026. eCollection 2022. Front Neurol. 2022. PMID: 35720065 Free PMC article.
-
Neurobiology of axonal transport defects in motor neuron diseases: Opportunities for translational research?Neurobiol Dis. 2017 Sep;105:283-299. doi: 10.1016/j.nbd.2017.02.004. Epub 2017 Feb 22. Neurobiol Dis. 2017. PMID: 28235672 Free PMC article. Review.
-
Microtubules, Membranes, and Movement: New Roles for Stathmin-2 in Axon Integrity.J Neurosci Res. 2024 Sep;102(9):e25382. doi: 10.1002/jnr.25382. J Neurosci Res. 2024. PMID: 39253877 Free PMC article. Review.
-
The Peripheral Nervous System in Amyotrophic Lateral Sclerosis: Opportunities for Translational Research.Front Neurosci. 2019 Jun 25;13:601. doi: 10.3389/fnins.2019.00601. eCollection 2019. Front Neurosci. 2019. PMID: 31293369 Free PMC article. Review.
-
Genome-wide RNA-seq of iPSC-derived motor neurons indicates selective cytoskeletal perturbation in Brown-Vialetto disease that is partially rescued by riboflavin.Sci Rep. 2017 Apr 6;7:46271. doi: 10.1038/srep46271. Sci Rep. 2017. PMID: 28382968 Free PMC article.
References
-
- Amin MR, Harris D, Cassel SG, Grimes E, Heiman‐Patterson T (2006) Sensory testing in the assessment of laryngeal sensation in patients with amyotrophic lateral sclerosis. Ann Otol Rhinol Laryngol 115:528–534. - PubMed
-
- Bartolini F, Tian G, Piehl M, Cassimeris L, Lewis SA, Cowan NJ (2005) Identification of a novel tubulin‐destabilizing protein related to the chaperone cofactor E. J Cell Sci 118:1197–1207. - PubMed
-
- Bellouze S, Schafer MK, Buttigieg D, Baillat G, Rabouille C, Haase G (2014) Golgi fragmentation in pmn mice is due to a defective ARF1/TBCE cross‐talk that coordinates COPI vesicle formation and tubulin polymerization. Hum Mol Genet 23:5961–5975. - PubMed
-
- Bento‐Abreu A, Van Damme P, Van Den Bosch L, Robberecht W (2010) The neurobiology of amyotrophic lateral sclerosis. Eur J Neurosci 31:2247–2265. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
