

Functional Heterogeneity in the Bed Nucleus of the Stria Terminalis
- PMID: 27488624
- PMCID: PMC4971356
- DOI: 10.1523/JNEUROSCI.0856-16.2016
Functional Heterogeneity in the Bed Nucleus of the Stria Terminalis
Abstract
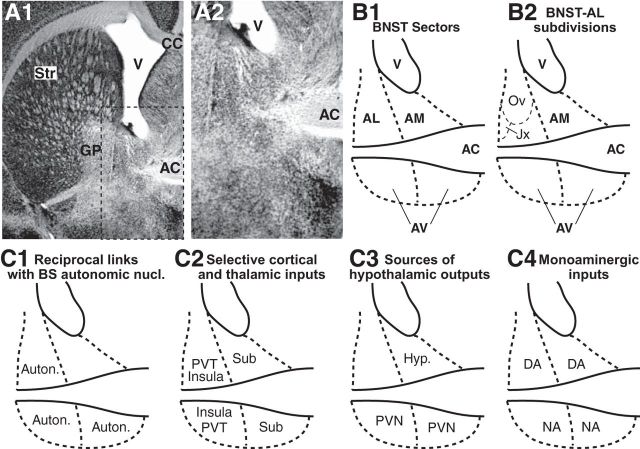
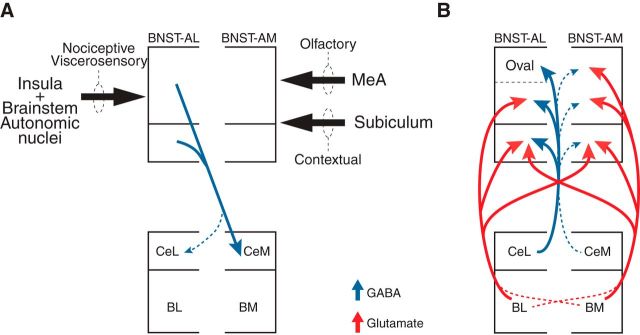
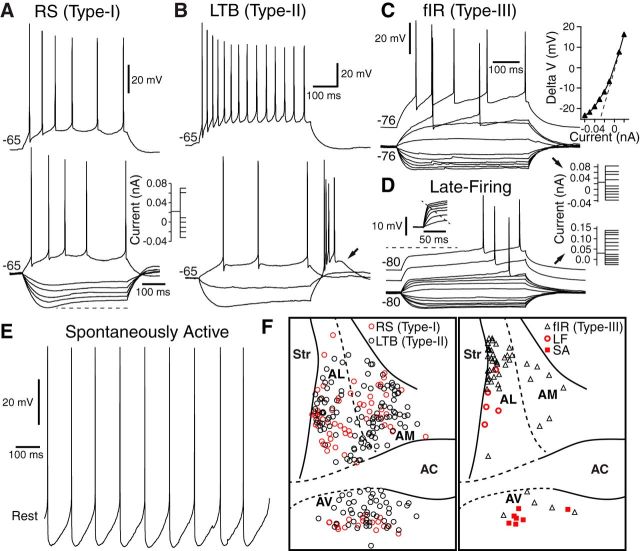
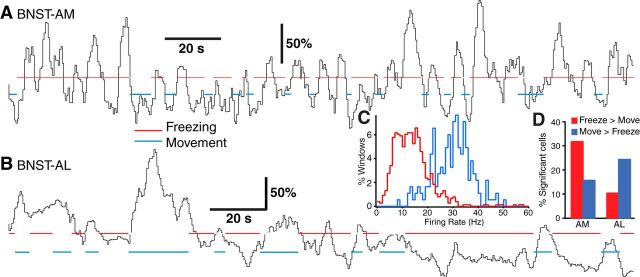
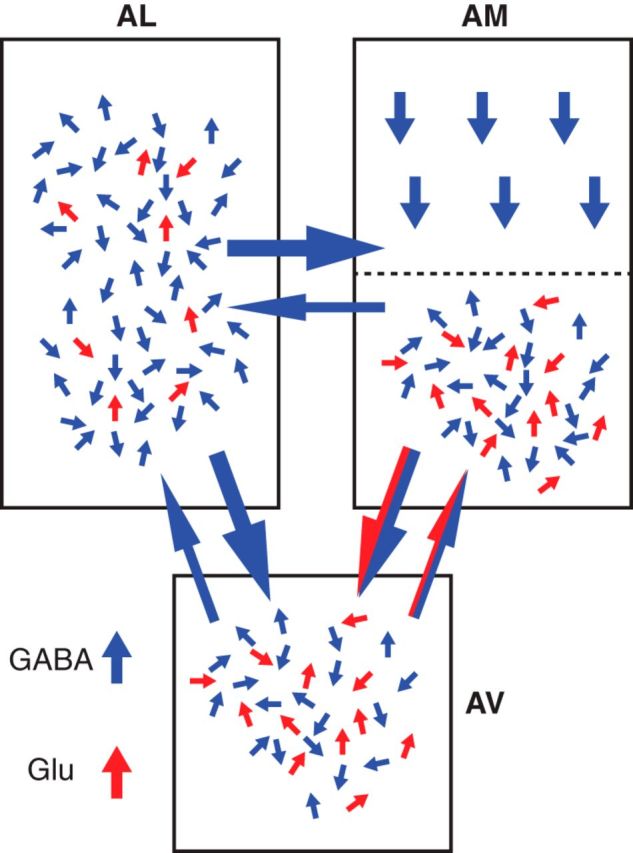
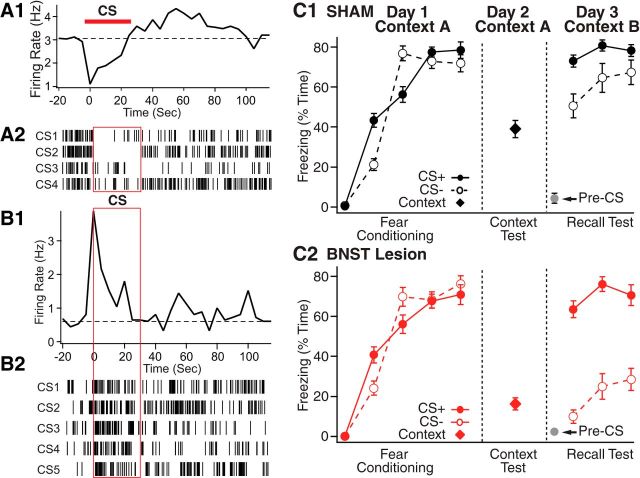
Early work stressed the differing involvement of the central amygdala (CeA) and bed nucleus of the stria terminalis (BNST) in the genesis of fear versus anxiety, respectively. In 2009, Walker, Miles, and Davis proposed a model of amygdala-BNST interactions to explain these functional differences. This model became extremely influential and now guides a new wave of studies on the role of BNST in humans. Here, we consider evidence for and against this model, in the process highlighting central principles of BNST organization. This analysis leads us to conclude that BNST's influence is not limited to the generation of anxiety-like responses to diffuse threats, but that it also shapes the impact of discrete threatening stimuli. It is likely that BNST-CeA interactions are involved in modulating responses to such threats. In addition, whereas current views emphasize the contributions of the anterolateral BNST region in anxiety, accumulating data indicate that the anteromedial and anteroventral regions also play a critical role. The presence of multiple functional subregions within the small volume of BNST raises significant technical obstacles for functional imaging studies in humans.
Keywords: BNST; amygdala; anxiety; fear.
Copyright © 2016 the authors 0270-6474/16/368038-12$15.00/0.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical