

Reciprocal Control of Drinking Behavior by Median Preoptic Neurons in Mice
- PMID: 27488641
- PMCID: PMC4971367
- DOI: 10.1523/JNEUROSCI.1244-16.2016
Reciprocal Control of Drinking Behavior by Median Preoptic Neurons in Mice
Abstract
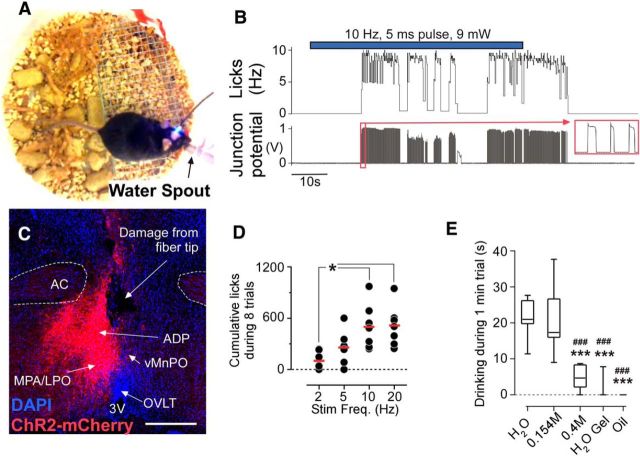
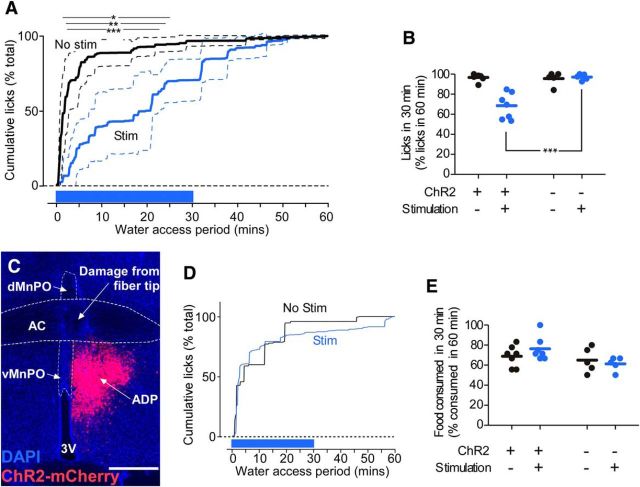
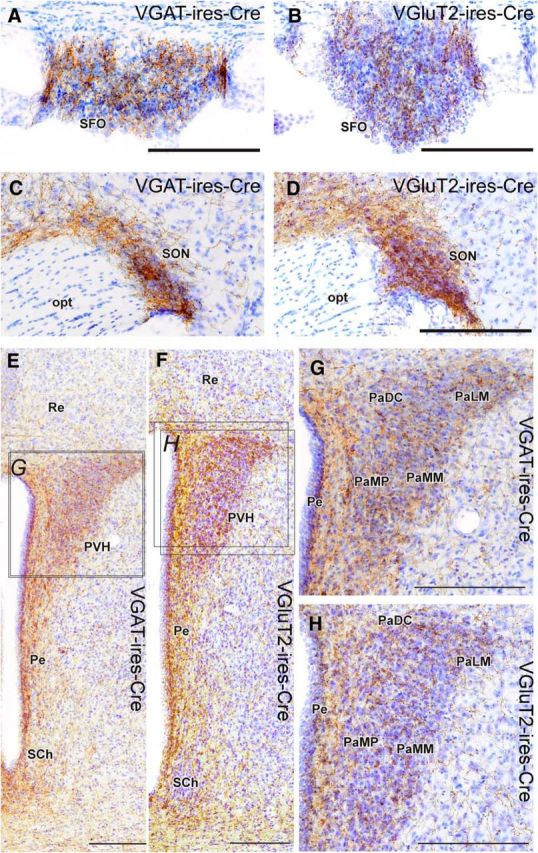
Stimulation of glutamatergic neurons in the subfornical organ drives drinking behavior, but the brain targets that mediate this response are not known. The densest target of subfornical axons is the anterior tip of the third ventricle, containing the median preoptic nucleus (MnPO) and organum vasculosum of the lamina terminalis (OVLT), a region that has also been implicated in fluid and electrolyte management. The neurochemical composition of this region is complex, containing both GABAergic and glutamatergic neurons, but the possible roles of these neurons in drinking responses have not been addressed. In mice, we show that optogenetic stimulation of glutamatergic neurons in MnPO/OVLT drives voracious water consumption, and that optogenetic stimulation of GABAergic neurons in the same region selectively reduces water consumption. Both populations of neurons have extensive projections to overlapping regions of the thalamus, hypothalamus, and hindbrain that are much more extensive than those from the subfornical organ, suggesting that the MnPO/OVLT serves as a key link in regulating drinking responses.
Significance statement: Neurons in the median preoptic nucleus (MnPO) and organum vasculosum of the lamina terminalis (OVLT) are known to regulate fluid/electrolyte homeostasis, but few studies have examined this issue with an appreciation for the neurochemical heterogeneity of these nuclei. Using Cre-Lox genetic targeting of Channelrhodospin-2 in transgenic mice, we demonstrate that glutamate and GABA neurons in the MnPO/OVLT reciprocally regulate water consumption. Stimulating glutamatergic MnPO/OVLT neurons induced water consumption, whereas stimulating GABAergic MnPO neurons caused a sustained and specific reduction in water consumption in dehydrated mice, the latter highlighting a heretofore unappreciated role of GABAergic MnPO neurons in thirst regulation. These observations represent an important advance in our understanding of the neural circuits involved in the regulation of fluid/electrolyte homeostasis.
Keywords: VGAT; VGLUT2; dehydration; lamina terminalis; optogenetics; thirst.
Copyright © 2016 the authors 0270-6474/16/368228-10$15.00/0.
Figures



References
-
- De Luca LA, Jr, Xu Z, Schoorlemmer GH, Thunhorst RL, Beltz TG, Menani JV, Johnson AK. Water deprivation-induced sodium appetite: humoral and cardiovascular mediators and immediate early genes. Am J Physiol Regul Integr Comp Physiol. 2002;282:R552–R559. doi: 10.1152/ajpregu.00295.2000. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases