

Review
doi: 10.1152/physiol.00006.2016.
Regulation of Gastrointestinal Smooth Muscle Function by Interstitial Cells
Affiliations
- PMID: 27488743
- PMCID: PMC5504460
- DOI: 10.1152/physiol.00006.2016
Item in Clipboard
Review
Regulation of Gastrointestinal Smooth Muscle Function by Interstitial Cells
Physiology (Bethesda).
2016 Sep.
Abstract
Interstitial cells of mesenchymal origin form gap junctions with smooth muscle cells in visceral smooth muscles and provide important regulatory functions. In gastrointestinal (GI) muscles, there are two distinct classes of interstitial cells, c-Kit(+) interstitial cells of Cajal and PDGFRα(+) cells, that regulate motility patterns. Loss of these cells may contribute to symptoms in GI motility disorders.
©2016 Int. Union Physiol. Sci./Am. Physiol. Soc.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the author(s).
Figures
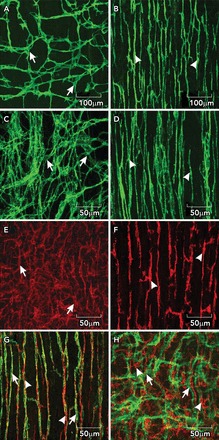
ICC and PDGFRα+ cells in the small intestine A and B: whole mounts of c-Kit+ ICC-MY (arrows) and ICC-DMP (arrowheads) in the murine small intestine. Note the well defined network structure of ICC-MY, and the more distributed and parallel orientation of ICC-DMP that run parallel to the circular muscle and are closely aligned with nerve processes of the deep muscular plexus. C and D: images from the small intestine of Macaca fascicularis (cynomolgus monkey) showing relatively the same distributions of ICC-MY (arrows) and ICC-DMP (arrowheads) in the primate intestine. E and F: distributions of PDGFRα+ cells at the level of the myenteric plexus (E; PDGFRα+-MY; arrows) and PDGFRα+ cells in the deep muscular plexus (F; PDGFRα+-DMP; arrowheads) in the murine intestine. Intramuscular ICC and PDGFRα+ are closely associated with enteric motor nerves. G: the relationship between ICC-DMP (green; arrows) in very close contact with inhibitory motor neurons highlighted by an antibody for nNOS (red; arrowheads). ICC and PDGFRα+ cells are distinct populations of interstitial cells. H: ICC-MY (green; arrows) and PDGFRα+ cells (red; arrowheads) in the same tissue. Scale bars are shown for each panel.
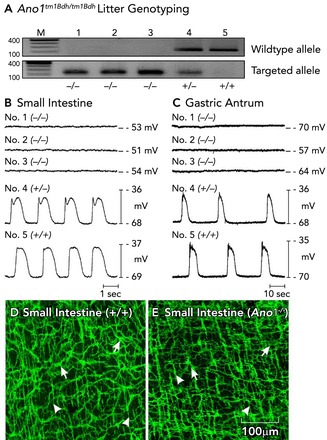
Loss of pacemaker activity in Ano1−/− mice A: genotyping of Ano1tm1Bdh/tm1bdh mice. The wild-type allele was absent in animals 1–3 in this representative litter, proving that these mice were Ano1−/−. Animals 4 and 5 were a heterozygote and a wild-type homozygote, respectively. B and C: intracellular electrical recordings from jejunal and antral circular muscles of each of the five animals. Slow waves were absent in the muscles of Ano1−/− mice. Slow waves with a normal amplitude and frequency were recorded from animals that were heterozygous or homozygous for Ano1 (animals 4 and 5, respectively). In contrast to what has been claimed by others (102), we observed no significant differences in the development or density of ICC-MY (pacemaker class of ICC in these organs; see Table 1). D and E: ICC [ICC-MY (arrows) and ICC-DMP (arrowheads)] have a normal distribution and density in the small intestines of Ano1−/− mice, suggesting that Ano1 is not critical for the development of ICC. Figure is redrawn from Ref. with permission. Scale bar for D and E is shown in E.
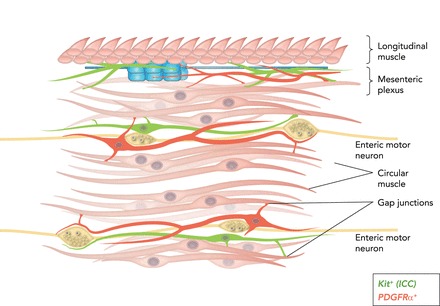
Cartoon showing SIP syncytium (94) c-Kit+ ICC (green) and PDGFRα+ (fibroblast-like cells; red) are intermixed with smooth muscle fibers in bundles of cells within the circular muscle layer (and in some regions in the longitudinal muscle layer as well). Intramuscular ICC and PDGFRα+ cells are closely associated with the processes of enteric motoneurons and express receptors, second-messenger pathways, and ion channels facilitating responses to enteric motor neurotransmitters. ICC and PDGFRα+ cells are electrically coupled to SMCs, so responses elicited in the interstitial cells can conduct to SMCs and regulate the excitability of the musculature. ICC-MY and PDGFRα+ cells are also found in the region of the myenteric plexus. ICC-MY are pacemaker cells and generate electrical slow waves. PDGFRα+ cells are responsive to purines. Here again, the interstitial cells are electrically coupled to circular and longitudinal SMCs, and slow waves conduct from ICC-MY to SMCs to generate a phasic pattern of contractions in the musculature.
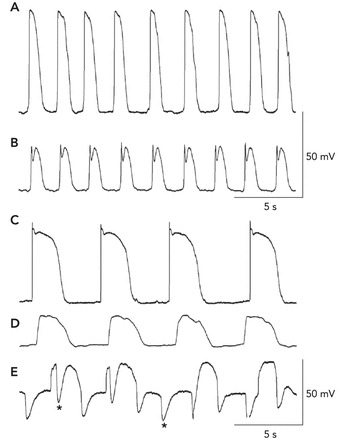
Recordings of electrical activity from the different cell types of the SIP syncytium A and B: slow waves recorded by impalements of an ICC-MY and a circular SMC in the murine small intestine (as in Refs. 60, 61), respectively. The resting potentials in these cells were −70 mV in the ICC-MY and −68 mV in the SMC. The slow-wave events in ICC are much larger in amplitude, and the upstroke depolarization is ∼1 V/s. Slow waves conduct to SMCs, and the smaller amplitude is a result of the passive nature of this communication (i.e., decremental conduction). C–E: recordings from an ICC-MY, a SMC, and a fibroblast-like cell of the rabbit small intestine (as in Refs. 58, 59), respectively. In these recordings, the resting potentials were −68 mV for the ICC-MY, −64 mV for the SMC, and −43 mV for the fibroblast-like cell. Note again that the amplitude and rise time of the slow waves recorded from ICC-MY are much greater than the events recorded in SMCs. Small oscillations at the frequency of slow waves were also observed in fibroblast-like cells, but a prominent activity in these cells was spontaneous transient hyperpolarizations (STHs; asterisks denote examples). These events appeared to be due to SK channels because they were blocked by apamin. STHs occurred during and between slow waves. Cells were identified by filling them with fluorescent dyes during impalements.
References
-
- Alberti E, Mikkelsen HB, Wang XY, Diaz M, Larsen JO, Huizinga JD, Jimenez M. Pacemaker activity and inhibitory neurotransmission in the colon of Ws/Ws mutant rats. Am J Physiol Gastrointest Liver Physiol 292: G1499–G1510, 2007. - PubMed
-
- Aranishi H, Kunisawa Y, Komuro T. Characterization of interstitial cells of Cajal in the subserosal layer of the guinea-pig colon. Cell Tiss Res 335: 323–329, 2009. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous