

NADPH Oxidases Are Essential for Macrophage Differentiation
- PMID: 27489105
- PMCID: PMC5025689
- DOI: 10.1074/jbc.M116.731216
NADPH Oxidases Are Essential for Macrophage Differentiation
Abstract
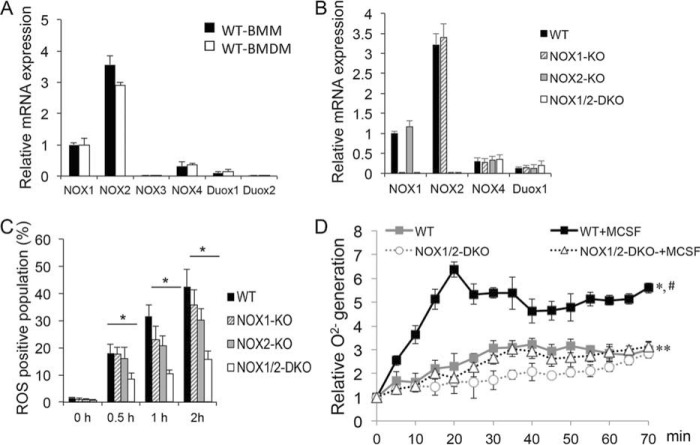
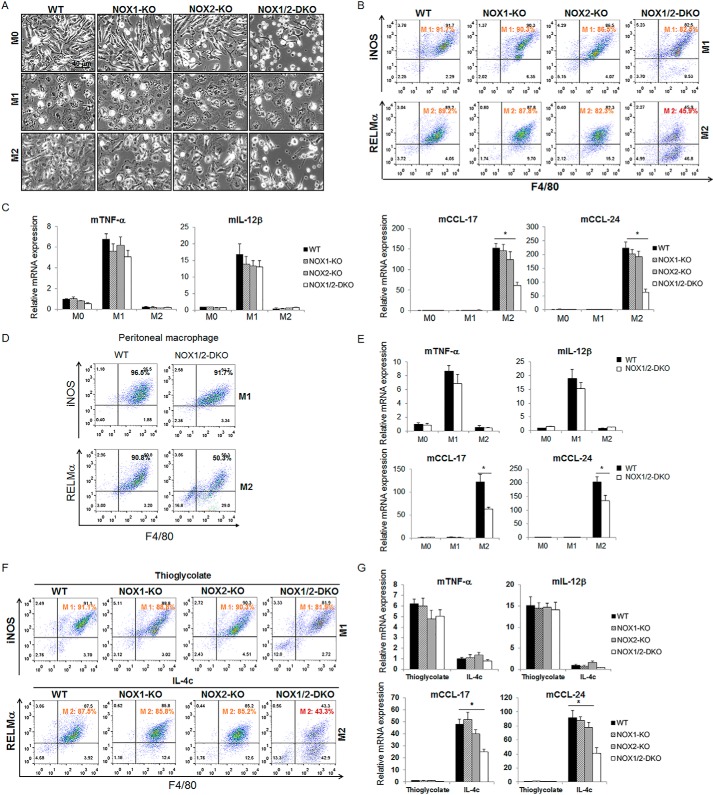
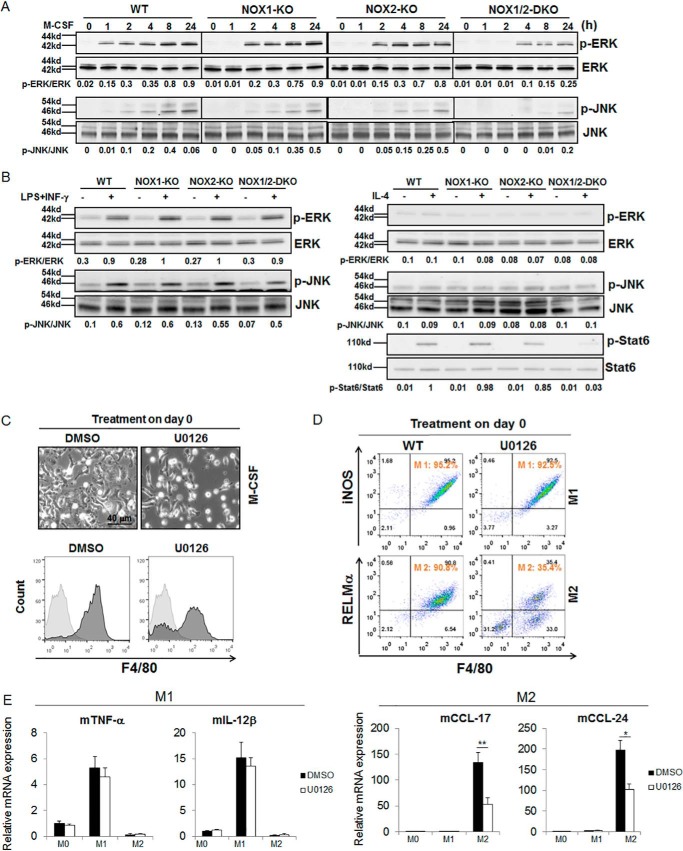
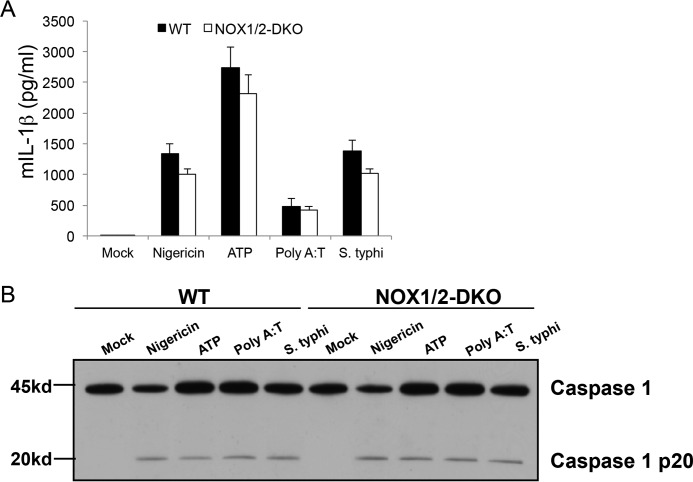
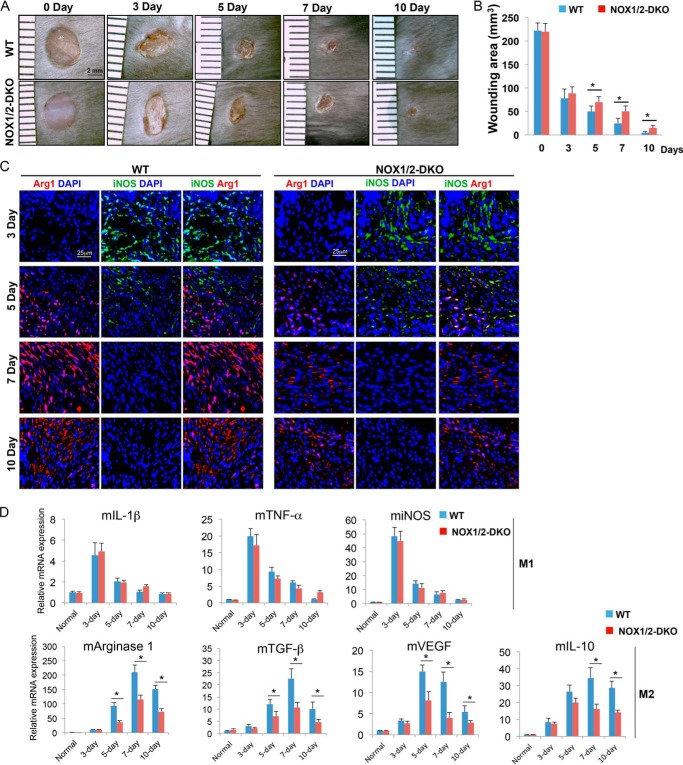
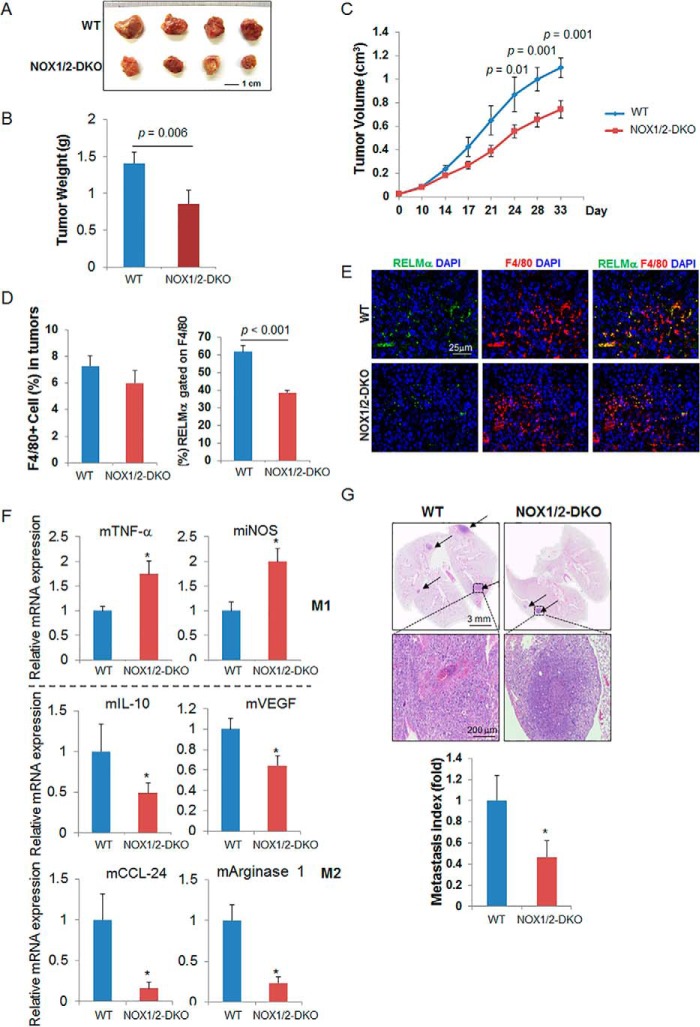
NADPH oxidases (NOXs) are involved in inflammation, angiogenesis, tumor growth, and osteoclast differentiation. However, the role of NOX1 and NOX2 in macrophage differentiation and tumor progression is still elusive. Here we report that NOX1 and NOX2 are critical for the differentiation of monocytes to macrophages, the polarization of M2-type but not M1-type macrophages, and the occurrence of tumor-associated macrophages (TAMs). We found that deletion of both NOX1 and NOX2 led to a dramatic decrease in ROS production in macrophages and resulted in impaired efficiency in monocyte-to-macrophage differentiation and M2-type macrophage polarization. We further showed that NOX1 and NOX2 were critical for the activation of the MAPKs JNK and ERK during macrophage differentiation and that the deficiency of JNK and ERK activation was responsible for the failure of monocyte-to-macrophage differentiation, in turn affecting M2 macrophage polarization. Furthermore, we demonstrated that the decrease in M2 macrophages and TAMs, concomitant with the reduction of cytokine and chemokine secretion, contributed to the delay in wound healing and the inhibition of tumor growth and metastasis in NOX1/2 double knockout mice compared with WT mice. Collectively, these data provide direct evidence that NOX1 and NOX2 deficiency impairs macrophage differentiation and the occurrence of M2-type TAMs during tumor development.
Keywords: NADPH oxidase; cell biology; cell differentiation; macrophage; reactive oxygen species (ROS).
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
Similar articles
-
Neurotoxic activation of microglia is promoted by a nox1-dependent NADPH oxidase.J Neurosci. 2008 Nov 12;28(46):12039-51. doi: 10.1523/JNEUROSCI.3568-08.2008. J Neurosci. 2008. PMID: 19005069 Free PMC article.
-
Role of NADPH oxidase-2 in lipopolysaccharide-induced matrix metalloproteinase expression and cell migration.Immunol Cell Biol. 2010 Feb;88(2):197-204. doi: 10.1038/icb.2009.87. Epub 2009 Nov 24. Immunol Cell Biol. 2010. PMID: 19935767
-
NADPH oxidase-derived reactive oxygen species are essential for differentiation of a mouse macrophage cell line (RAW264.7) into osteoclasts.J Med Invest. 2009 Feb;56(1-2):33-41. doi: 10.2152/jmi.56.33. J Med Invest. 2009. PMID: 19262012
-
Combating oxidative stress in vascular disease: NADPH oxidases as therapeutic targets.Nat Rev Drug Discov. 2011 Jun;10(6):453-71. doi: 10.1038/nrd3403. Nat Rev Drug Discov. 2011. PMID: 21629295 Free PMC article. Review.
-
The many roles of NOX2 NADPH oxidase-derived ROS in immunity.Semin Immunopathol. 2010 Dec;32(4):415-30. doi: 10.1007/s00281-010-0221-0. Epub 2010 Aug 28. Semin Immunopathol. 2010. PMID: 20803017 Review.
Cited by
-
Circadian clock protein BMAL1 regulates IL-1β in macrophages via NRF2.Proc Natl Acad Sci U S A. 2018 Sep 4;115(36):E8460-E8468. doi: 10.1073/pnas.1800431115. Epub 2018 Aug 20. Proc Natl Acad Sci U S A. 2018. PMID: 30127006 Free PMC article.
-
Macrophage Polarization and Reprogramming in Acute Inflammation: A Redox Perspective.Antioxidants (Basel). 2022 Jul 19;11(7):1394. doi: 10.3390/antiox11071394. Antioxidants (Basel). 2022. PMID: 35883885 Free PMC article. Review.
-
Immunometabolism changes in fibrosis: from mechanisms to therapeutic strategies.Front Pharmacol. 2023 Jul 27;14:1243675. doi: 10.3389/fphar.2023.1243675. eCollection 2023. Front Pharmacol. 2023. PMID: 37576819 Free PMC article. Review.
-
Distinct immune signatures discriminate between asymptomatic and presymptomatic SARS-CoV-2pos subjects.Cell Res. 2021 Nov;31(11):1148-1162. doi: 10.1038/s41422-021-00562-1. Epub 2021 Sep 24. Cell Res. 2021. PMID: 34561618 Free PMC article.
-
The pro-inflammatory signature of lipopolysaccharide in spontaneous contracting embryoid bodies differentiated from mouse embryonic stem cells.J Cell Mol Med. 2023 Jul;27(14):2045-2058. doi: 10.1111/jcmm.17805. Epub 2023 Jun 14. J Cell Mol Med. 2023. PMID: 37315183 Free PMC article.
References
-
- Jiang F., Zhang Y., and Dusting G. J. (2011) NADPH oxidase-mediated redox signaling: roles in cellular stress response, stress tolerance, and tissue repair. Pharmacol. Rev. 63, 218–242 - PubMed
-
- Chan E. C., Jiang F., Peshavariya H. M., and Dusting G. J. (2009) Regulation of cell proliferation by NADPH oxidase-mediated signaling: potential roles in tissue repair, regenerative medicine and tissue engineering. Pharmacol. Ther. 122, 97–108 - PubMed
-
- Bedard K., and Krause K. H. (2007) The NOX family of ROS-generating NADPH oxidases: physiology and pathophysiology. Physiol. Rev. 87, 245–313 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
