

Identification and characterization of wheat drought-responsive MYB transcription factors involved in the regulation of cuticle biosynthesis
- PMID: 27489236
- PMCID: PMC5049387
- DOI: 10.1093/jxb/erw298
Identification and characterization of wheat drought-responsive MYB transcription factors involved in the regulation of cuticle biosynthesis
Abstract
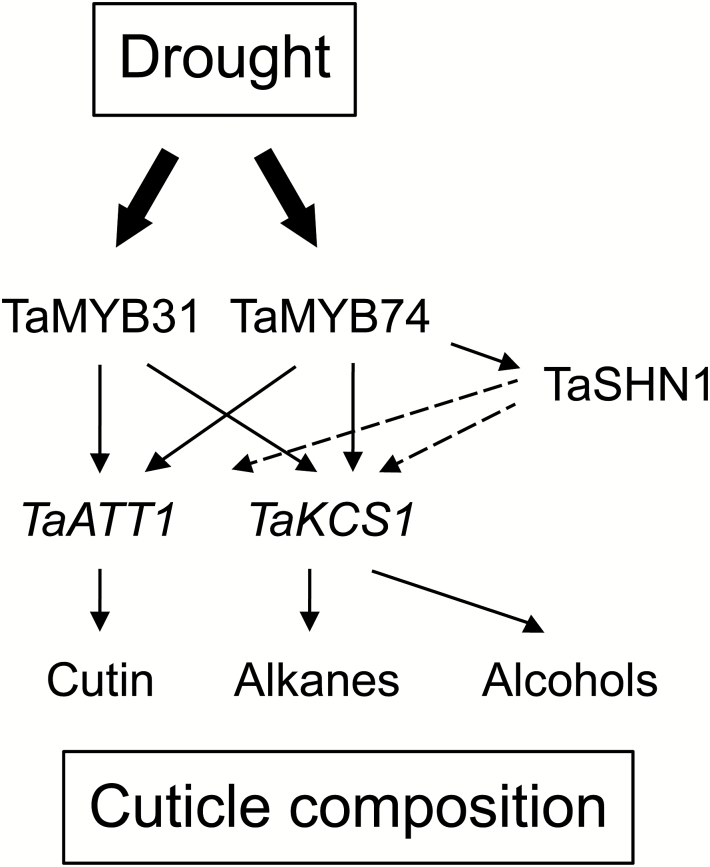
A plant cuticle forms a hydrophobic layer covering plant organs, and plays an important role in plant development and protection from environmental stresses. We examined epicuticular structure, composition, and a MYB-based regulatory network in two Australian wheat cultivars, RAC875 and Kukri, with contrasting cuticle appearance (glaucousness) and drought tolerance. Metabolomics and microscopic analyses of epicuticular waxes revealed that the content of β-diketones was the major compositional and structural difference between RAC875 and Kukri. The content of β-diketones remained the same while those of alkanes and primary alcohols were increased by drought in both cultivars, suggesting that the interplay of all components rather than a single one defines the difference in drought tolerance between cultivars. Six wheat genes encoding MYB transcription factors (TFs) were cloned; four of them were regulated in flag leaves of both cultivars by rapid dehydration and/or slowly developing cyclic drought. The involvement of selected MYB TFs in the regulation of cuticle biosynthesis was confirmed by a transient expression assay in wheat cell culture, using the promoters of wheat genes encoding cuticle biosynthesis-related enzymes and the SHINE1 (SHN1) TF. Two functional MYB-responsive elements, specifically recognized by TaMYB74 but not by other MYB TFs, were localized in the TdSHN1 promoter. Protein structural determinants underlying the binding specificity of TaMYB74 for functional DNA cis-elements were defined, using 3D protein molecular modelling. A scheme, linking drought-induced expression of the investigated TFs with downstream genes that participate in the synthesis of cuticle components, is proposed.
Keywords: Abiotic stress; MYB and SHINE1 transcription factors; cuticle; drought; molecular model; water deficit; wax; wheat.; β-diketone.
© The Author 2016. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures









References
-
- Adamski NM, Bush MS, Simmonds J, Turner AS, Mugford SG, Jones A, Findlay K, Pedentchouk N, Wettstein-Knowles P, Uauy C. 2013. The Inhibitor of wax 1 locus (Iw1) prevents formation of β- and OH-β-diketones in wheat cuticular waxes and maps to a sub-cM interval on chromosome arm 2BS. The Plant Journal 74, 989–1002. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
