

Structures of TorsinA and its disease-mutant complexed with an activator reveal the molecular basis for primary dystonia
- PMID: 27490483
- PMCID: PMC4999309
- DOI: 10.7554/eLife.17983
Structures of TorsinA and its disease-mutant complexed with an activator reveal the molecular basis for primary dystonia
Abstract
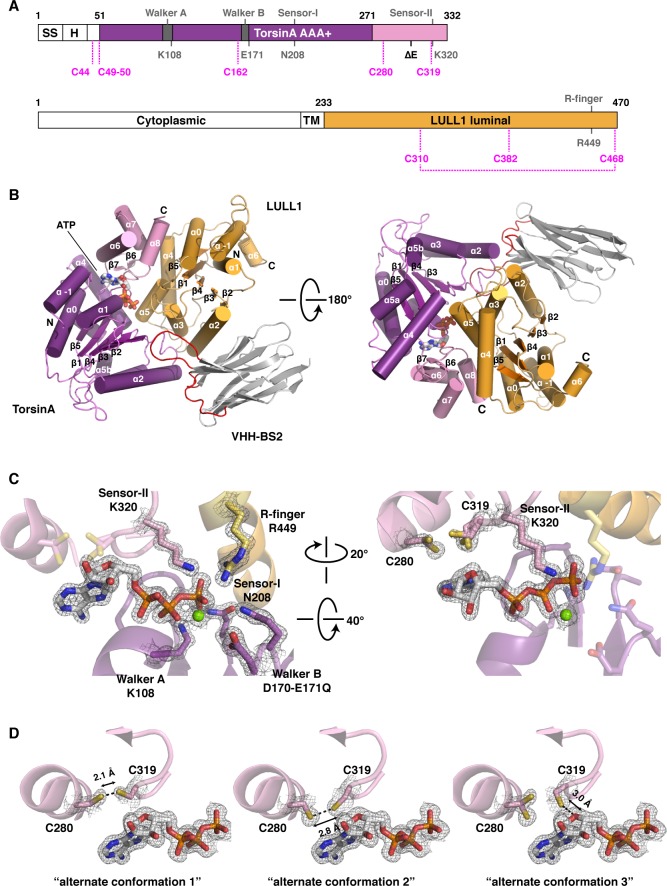
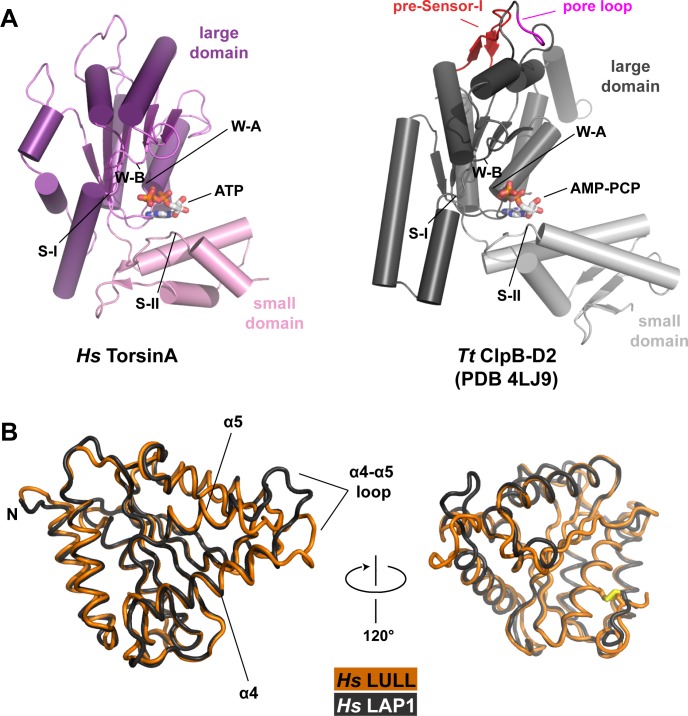
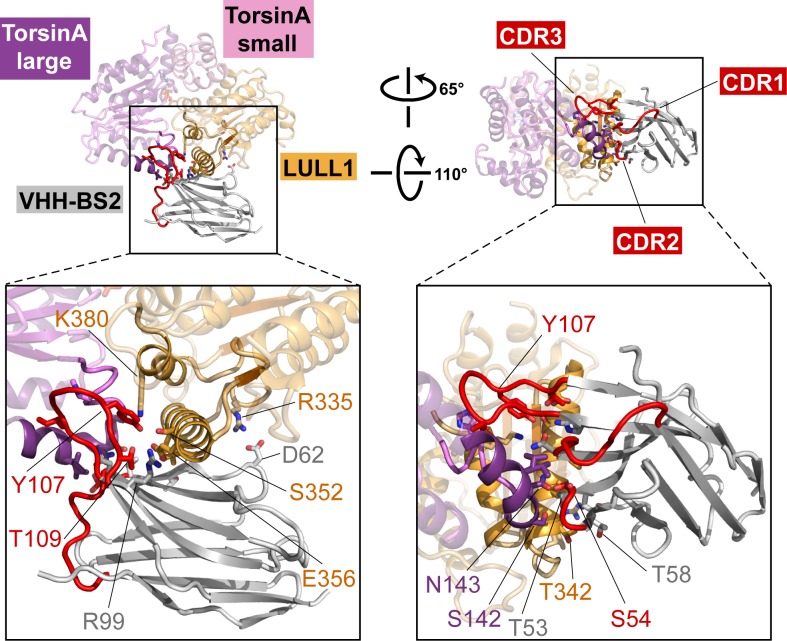
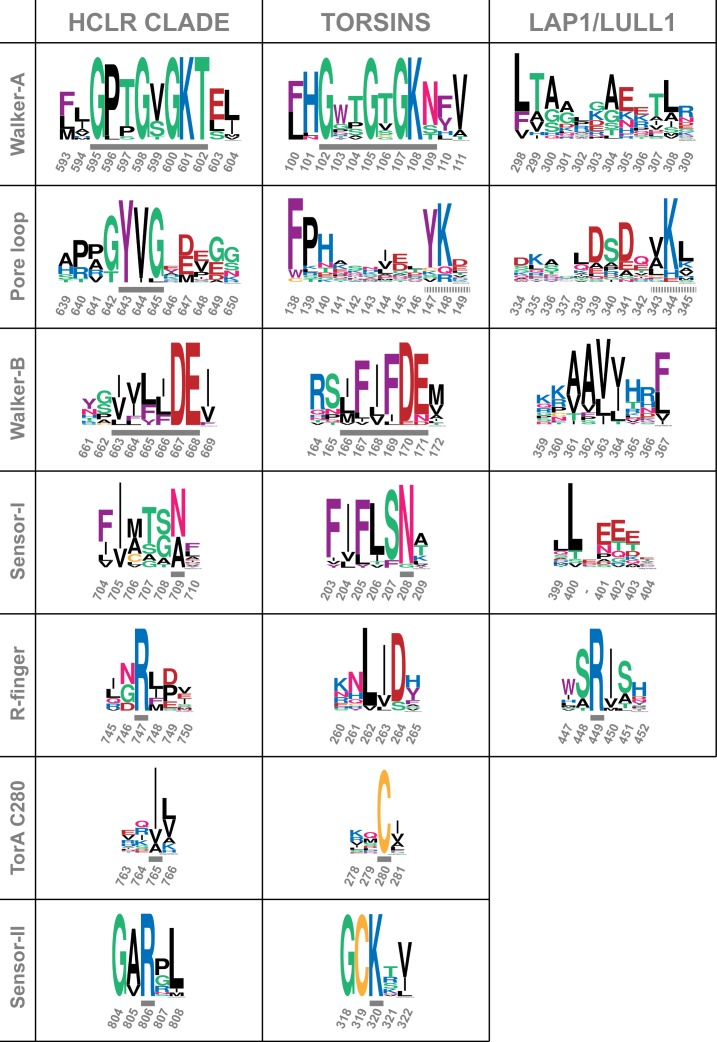
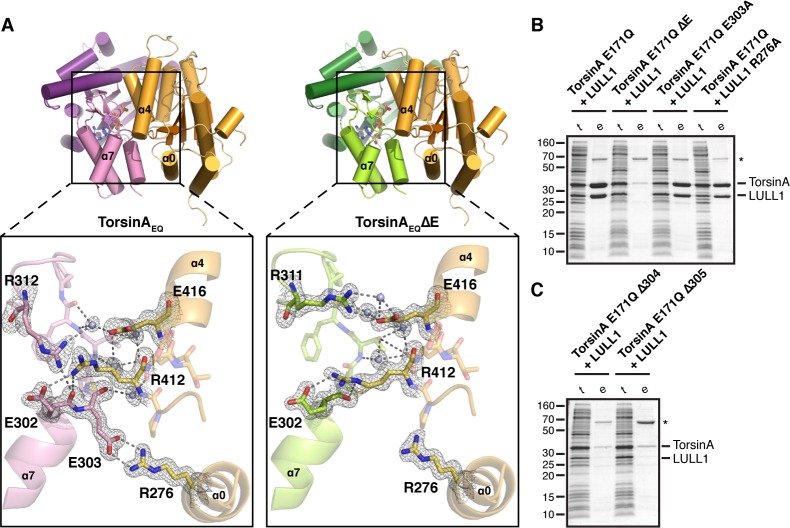
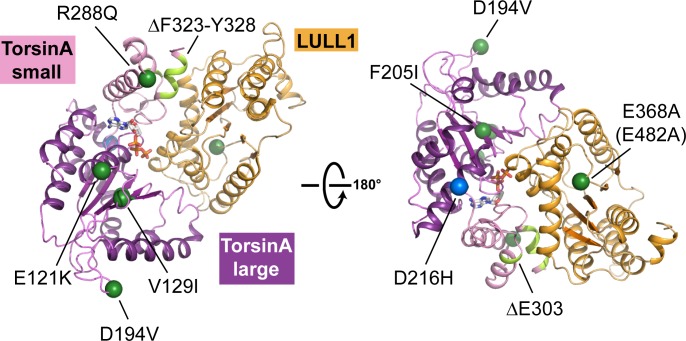
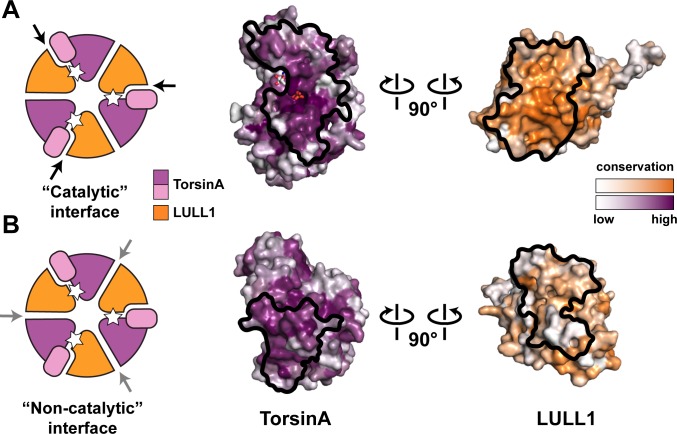
The most common cause of early onset primary dystonia, a neuromuscular disease, is a glutamate deletion (ΔE) at position 302/303 of TorsinA, a AAA+ ATPase that resides in the endoplasmic reticulum. While the function of TorsinA remains elusive, the ΔE mutation is known to diminish binding of two TorsinA ATPase activators: lamina-associated protein 1 (LAP1) and its paralog, luminal domain like LAP1 (LULL1). Using a nanobody as a crystallization chaperone, we obtained a 1.4 Å crystal structure of human TorsinA in complex with LULL1. This nanobody likewise stabilized the weakened TorsinAΔE-LULL1 interaction, which enabled us to solve its structure at 1.4 Å also. A comparison of these structures shows, in atomic detail, the subtle differences in activator interactions that separate the healthy from the diseased state. This information may provide a structural platform for drug development, as a small molecule that rescues TorsinAΔE could serve as a cure for primary dystonia.
Keywords: AAA+ ATPases; biophysics; cell biology; dystonia; human; nuclear envelope; structural biology.
Conflict of interest statement
FED, BAS and TUS: Filed a provisional patent application protecting the use of the crystal structures (U.S.P.T.O. No. 62/330,683). The other authors declare that no competing interests exist.
Figures


References
-
- Adams PD, Afonine PV, Bunkóczi G, Chen VB, Davis IW, Echols N, Headd JJ, Hung LW, Kapral GJ, Grosse-Kunstleve RW, McCoy AJ, Moriarty NW, Oeffner R, Read RJ, Richardson DC, Richardson JS, Terwilliger TC, Zwart PH. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallographica Section D Biological Crystallography. 2010;66:213–221. doi: 10.1107/S0907444909052925. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
