

Two-photon excited fluorescence of intrinsic fluorophores enables label-free assessment of adipose tissue function
- PMID: 27491409
- PMCID: PMC4974509
- DOI: 10.1038/srep31012
Two-photon excited fluorescence of intrinsic fluorophores enables label-free assessment of adipose tissue function
Abstract
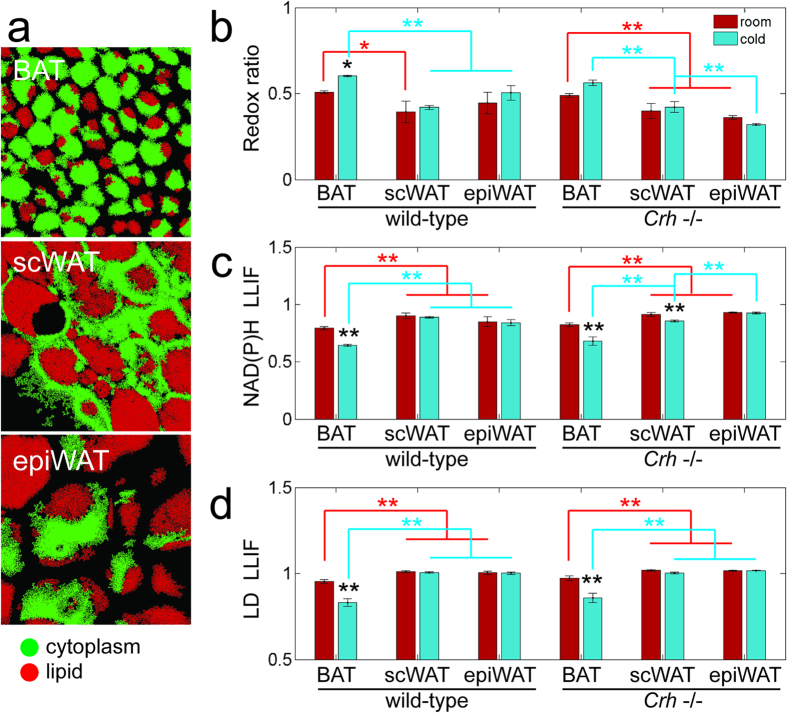
Current methods for evaluating adipose tissue function are destructive or have low spatial resolution. These limit our ability to assess dynamic changes and heterogeneous responses that occur in healthy or diseased subjects, or during treatment. Here, we demonstrate that intrinsic two-photon excited fluorescence enables functional imaging of adipocyte metabolism with subcellular resolution. Steady-state and time-resolved fluorescence from intracellular metabolic co-factors and lipid droplets can distinguish the functional states of excised white, brown, and cold-induced beige fat. Similar optical changes are identified when white and brown fat are assessed in vivo. Therefore, these studies establish the potential of non-invasive, high resolution, endogenous contrast, two-photon imaging to identify distinct adipose tissue types, monitor their functional state, and characterize heterogeneity of induced responses.
Figures






References
-
- Alberti K. G. et al. Harmonizing the metabolic syndrome: a joint interim statement of the International Diabetes Federation Task Force on Epidemiology and Prevention; National Heart, Lung, and Blood Institute; American Heart Association; World Heart Federation; International Atherosclerosis Society; and International Association for the Study of Obesity. Circulation 120, 1640–1645, doi: 10.1161/CIRCULATIONAHA.109.192644 (2009). - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
