

Coiled-coils: The long and short of it
- PMID: 27492088
- PMCID: PMC5082667
- DOI: 10.1002/bies.201600062
Coiled-coils: The long and short of it
Abstract
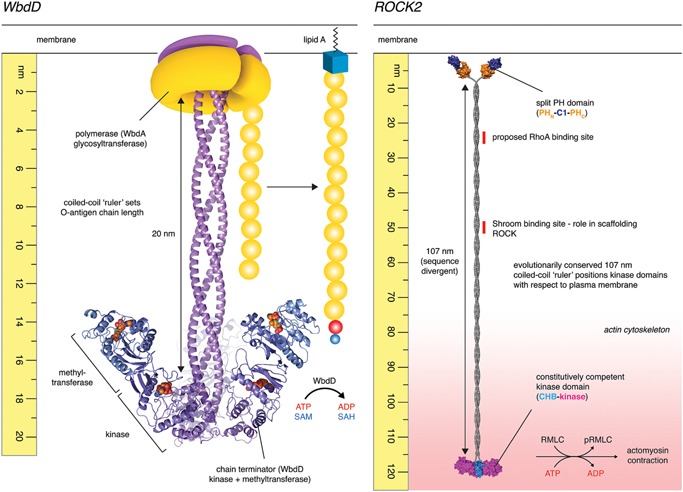
Coiled-coils are found in proteins throughout all three kingdoms of life. Coiled-coil domains of some proteins are almost invariant in sequence and length, betraying a structural and functional role for amino acids along the entire length of the coiled-coil. Other coiled-coils are divergent in sequence, but conserved in length, thereby functioning as molecular spacers. In this capacity, coiled-coil proteins influence the architecture of organelles such as centrioles and the Golgi, as well as permit the tethering of transport vesicles. Specialized coiled-coils, such as those found in motor proteins, are capable of propagating conformational changes along their length that regulate cargo binding and motor processivity. Coiled-coil domains have also been identified in enzymes, where they function as molecular rulers, positioning catalytic activities at fixed distances. Finally, while coiled-coils have been extensively discussed for their potential to nucleate and scaffold large macromolecular complexes, structural evidence to substantiate this claim is relatively scarce.
Keywords: allostery; coiled-coil; molecular ruler; molecular spacer; scaffold.
© 2016 The Authors BioEssays Published by WILEY Periodicals, Inc.
Figures
Similar articles
-
Coiled-coil protein composition of 22 proteomes--differences and common themes in subcellular infrastructure and traffic control.BMC Evol Biol. 2005 Nov 16;5:66. doi: 10.1186/1471-2148-5-66. BMC Evol Biol. 2005. PMID: 16288662 Free PMC article.
-
The coiled coils of cohesin are conserved in animals, but not in yeast.PLoS One. 2009;4(3):e4674. doi: 10.1371/journal.pone.0004674. Epub 2009 Mar 5. PLoS One. 2009. PMID: 19262687 Free PMC article.
-
Sequence divergence of coiled coils--structural rods, myosin filament packing, and the extraordinary conservation of cohesins.J Struct Biol. 2006 May;154(2):111-21. doi: 10.1016/j.jsb.2006.01.001. Epub 2006 Feb 3. J Struct Biol. 2006. PMID: 16495084
-
Transport Vesicle Tethering at the Trans Golgi Network: Coiled Coil Proteins in Action.Front Cell Dev Biol. 2016 Mar 15;4:18. doi: 10.3389/fcell.2016.00018. eCollection 2016. Front Cell Dev Biol. 2016. PMID: 27014693 Free PMC article. Review.
-
Pharmacological interference with protein-protein interactions mediated by coiled-coil motifs.Handb Exp Pharmacol. 2008;(186):461-82. doi: 10.1007/978-3-540-72843-6_19. Handb Exp Pharmacol. 2008. PMID: 18491064 Review.
Cited by
-
Mud binds the kinesin-14 Ncd in Drosophila.Biochem Biophys Rep. 2021 May 13;26:101016. doi: 10.1016/j.bbrep.2021.101016. eCollection 2021 Jul. Biochem Biophys Rep. 2021. PMID: 34027137 Free PMC article.
-
Molecular mechanism of Afadin substrate recruitment to the receptor phosphatase PTPRK via its pseudophosphatase domain.Elife. 2022 Oct 20;11:e79855. doi: 10.7554/eLife.79855. Elife. 2022. PMID: 36264065 Free PMC article.
-
Tunable Protein Hydrogels: Present State and Emerging Development.Adv Biochem Eng Biotechnol. 2021;178:63-97. doi: 10.1007/10_2021_167. Adv Biochem Eng Biotechnol. 2021. PMID: 33860358
-
Structural Perspectives on the Mechanism of Soluble Guanylate Cyclase Activation.Int J Mol Sci. 2021 May 21;22(11):5439. doi: 10.3390/ijms22115439. Int J Mol Sci. 2021. PMID: 34064029 Free PMC article. Review.
-
TipN's involvement with centromere segregation in Caulobacter crescentus.bioRxiv [Preprint]. 2023 Dec 21:2023.12.20.572679. doi: 10.1101/2023.12.20.572679. bioRxiv. 2023. Update in: Mol Biol Cell. 2024 May 1;35(5):ar68. doi: 10.1091/mbc.E23-12-0503. PMID: 38187783 Free PMC article. Updated. Preprint.
References
-
- Crick FH. 1952. Is alpha‐keratin a coiled coil? Nature 170: 882–3. - PubMed
-
- Wilson IA, Skehel JJ, Wiley DC. 1981. Structure of the haemagglutinin membrane glycoprotein of influenza virus at 3A resolution. Nature 289: 366–73. - PubMed
-
- Crick FH. 1953. The Fourier transform of a coiled‐coil. Acta Crystallogr 6: 685–9.
-
- Crick FH. 1953. The packing of α‐helices: simple coiled‐coils. Acta Crystallogr 6: 689–97.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
