

Review
doi: 10.1038/nrc.2016.71.
Epub 2016 Jul 29.
From Krebs to clinic: glutamine metabolism to cancer therapy
Affiliations
- PMID: 27492215
- PMCID: PMC5484415
- DOI: 10.1038/nrc.2016.71
Item in Clipboard
Review
From Krebs to clinic: glutamine metabolism to cancer therapy
Nat Rev Cancer.
2016 Oct.
Erratum in
-
From Krebs to clinic: glutamine metabolism to cancer therapy.Nat Rev Cancer. 2016 Dec;16(12):773. doi: 10.1038/nrc.2016.131. Epub 2016 Nov 11. Nat Rev Cancer. 2016. PMID: 28704359 No abstract available.
-
From Krebs to clinic: glutamine metabolism to cancer therapy.Nat Rev Cancer. 2016 Nov;16(11):749. doi: 10.1038/nrc.2016.114. Epub 2016 Oct 14. Nat Rev Cancer. 2016. PMID: 28704361 No abstract available.
Abstract
The resurgence of research into cancer metabolism has recently broadened interests beyond glucose and the Warburg effect to other nutrients, including glutamine. Because oncogenic alterations of metabolism render cancer cells addicted to nutrients, pathways involved in glycolysis or glutaminolysis could be exploited for therapeutic purposes. In this Review, we provide an updated overview of glutamine metabolism and its involvement in tumorigenesis in vitro and in vivo, and explore the recent potential applications of basic science discoveries in the clinical setting.
Conflict of interest statement
There is NO competing interest.
Figures

Timeline of key discoveries in mammalian glutamine metabolism and cancer α-KG, α-ketoglutarate; GLUD, glutamate dehydrogenase.
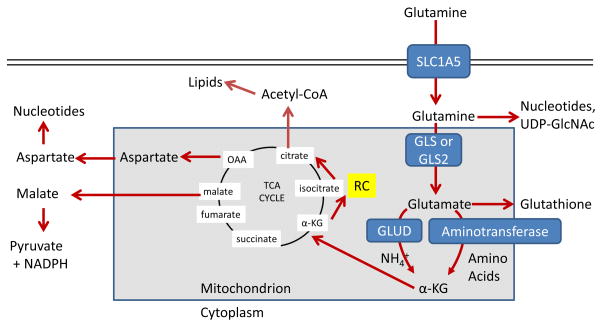
Glutamine enters the mammalian cell through transporters such as SLC1A5 (also
known as ASCT2) . Glutamine
itself can contribute to nucleotide biosynthesis and uridine diphosphate
N-acetylglucosamine (UDP-GlcNAc) synthesis for support of protein folding and
trafficking , or is
converted to glutamate by glutaminase (GLS or GLS2) . Glutamate can contribute to the
synthesis of glutathione ,
and has many other metabolic fates in the cell that impact on several inborn
errors of metabolism, which were recently reviewed . Glutamate is converted to
α-ketoglutarate (αKG) through one of two sets of enzymes,
glutamate dehydrogenase (GLUD1 or GLUD2, henceforth referred to collectively as
GLUD) or aminotransferases .
While the byproduct of GLUD is NH4+, the byproduct
of aminotransferase reactions is other amino acids. Note that aminotransferases
may be present either in the cytoplasm or the mitochondria.
α-ketoglutarate enters the tricarboxylic acid (TCA) cycle and can
provide energy for the cell. Malate exiting the TCA cycle can produce pyruvate
and NADPH for reducing equivalents , and oxaloacetate (OAA) can be converted to aspartate
to support nucleotide synthesis . These two pathways are illustrated in more detail in Figure 4. Alternately, α-KG can
proceed backwards through the TCA cycle, in a process called reductive
carboxylation (RC) to produce citrate, which supports synthesis of acetyl-CoA
and lipids .
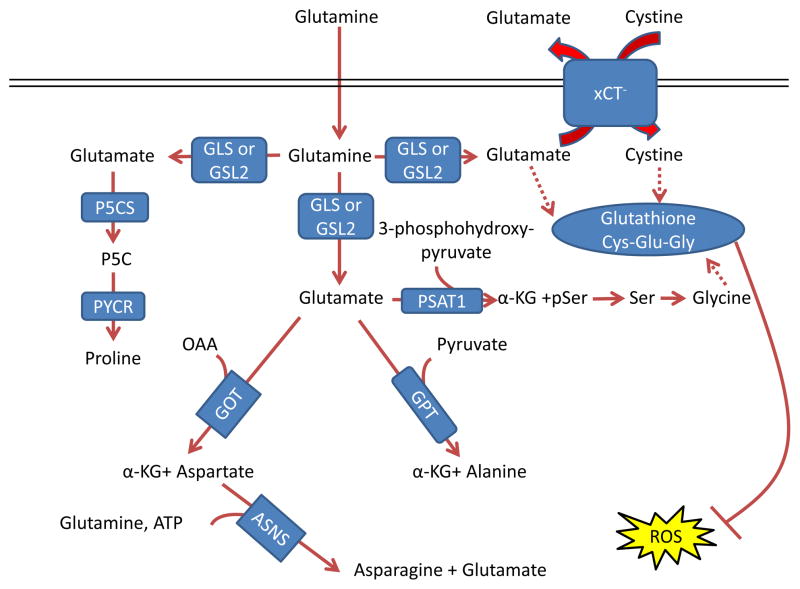
Glutamate acts as a nitrogen donor for the transamination involved in the
production of ‘dispensable amino acids’ alanine, aspartate, and
serine through the actions of glutamic-oxaloacetic transaminase (GOT), glutamic
pyruvate transaminase (GPT) and phosphoserine aminotransferase 1 (PSAT1),
respectively. Glutamine can also act as a nitrogen donor for asparagine through
asparagine synthetase (ASNS). In a reaction independent of transamination,
proline can be synthesized by conversion of glutamate to pyrroline-5-carboxylate
(P5C) by pyrroline-5-carboxylate synthase (P5CS; also known as aldehyde
dehydrogenase 18 family member A1, (ALDH18A1)) and subsequently to proline by
pyrroline-5-carboxylate reductase 1 (PYCR1) and PYCR2. Glutamine also
contributes to the tripeptide glutathione (composed of glutamate, cysteine and
glycine), which neutralizes the ROS H2O2
. The first step in
glutathione synthesis is the condensation of glutamate and cysteine through
glutamate-cysteine ligase (GCL; not shown in the figure). Glutamine input
directly contributes to the availability of cysteine and glycine for production
of glutathione. Glutamate can be exchanged for cystine (which is quickly reduced
to cysteine inside the cell) through the xCT antiporter (a heterodimer of
SLC7A11 and SCL3A2), which has been shown to be important in a variety of
cancers and has been considered as a drug target , . Glycine is next added by glutathione synthetase (GSS; not
shown in the figure). Additionally, glutamate can contribute to glycine through
transamination by PSAT1 into phosphoserine (pSer) and α-ketoglutarate
(αKG) and subsequent conversion to glycine through serine
hydroxymethyltransferase (SHMT; not shown in the figure) as part of the
one-carbon metabolism pathway, which has been shown in numerous studies to be
critical in cancer metabolism and is also reviewed in this Focus Issue by Dr.
Karen Vousden , , . GLS, kidney-type glutaminase; GLS2, liver-type
glutaminase; GLUD, glutamate dehydrogenase; OAA, oxaloacetate.
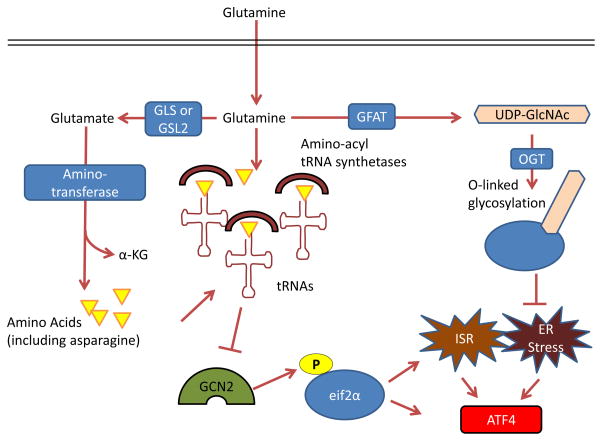
GCN2, a serine-threonine kinase with a regulatory domain that is structurally
similar to histidine-tRNA synthetase, is allosterically activated by uncharged
tRNAs with amino acid deprivation (including glutamine deprivation) and in turn
activates the integrated stress response (ISR) , ,
. Glutamine can
suppress GCN2 activation through its contribution to amino acid pools by
aminotransferases , –. To control endoplasmic reticulum (ER)
homeostasis, glutamine supports protein folding and trafficking through its
contribution to uridine diphosphate N-acetylglucosamine (UDP-GlcNAc) as part of
the hexosamine biosynthesis pathway. Glutamine is the substrate for glutamine
fructose-6-phosphate aminotransferase (GFAT), which is the key rate-limiting
enzyme in the hexosamine pathway, and the downstream product UDP-GlcNAc is a
substrate for O-linked glycosylation through O-linked
β-N-acetylglucosamine transferase (OGT). Thus, glutamine deprivation can
lead to improper protein folding and chaperoning and ER stress . A key output of both the
ISR and of ER stress is activating transcription factor 4 (ATF4), which is
induced via cap-independent translation downstream of eukaryotic translation
initiation factor 2α (eIF2α) phosphorylation by GCN2 or other
kinases . α-KG,
α-ketoglutarate; GLS, kidney-type glutaminase; GLS2, liver-type
glutaminase.
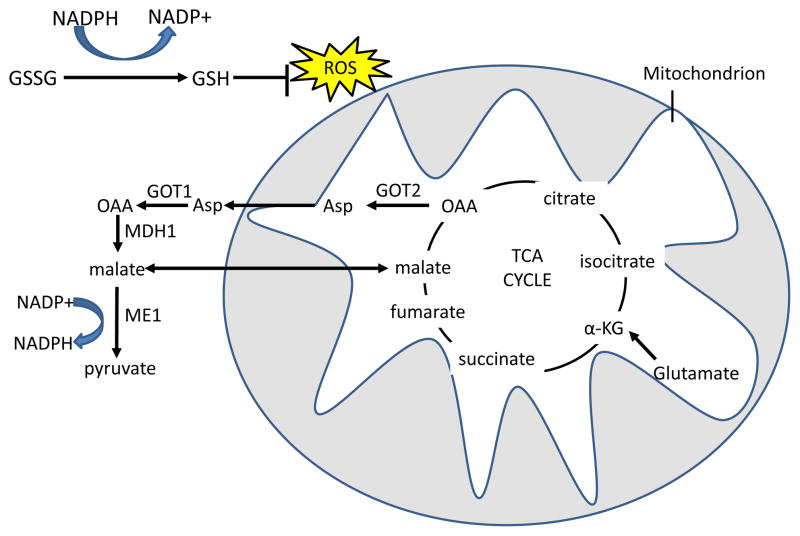
Reduced glutathione (GSH) neutralizes H2O2 with the
glutathione peroxidase enzyme, and oxidized glutathione (GSSG) is reduced by
NADPH and glutathione reductase to regenerate GSH. In the first pathway,
glutamine-derived malate is transported out of the mitochondria, and is
converted by malic enzyme 1 (ME1) to pyruvate, reducing one molecule of
NADP+ to NADPH. In the malate-aspartate shuttle-related second pathway,
found in mutant KRAS-transformed cells, aspartate that is produced from GOT2
mediated transamination of glutamine-derived oxaloacetate (OAA) is transported
out of the mitochondria. Aspartate is then converted in the cytosol back to OAA
by GOT1 and then to malate by malate dehydrogenase 1 (MDH1), which is in turn
processed to pyruvate by ME1 to produce one molecule of NADPH . The fate of glutamine-derived
pyruvate is similar to glucose-derived pyruvate in that much of it is expelled
as lactate. α-KG,
α-ketoglutarate; TCA, tricarboxylic acid; GOT, glutamic oxaloacetate
transaminase.
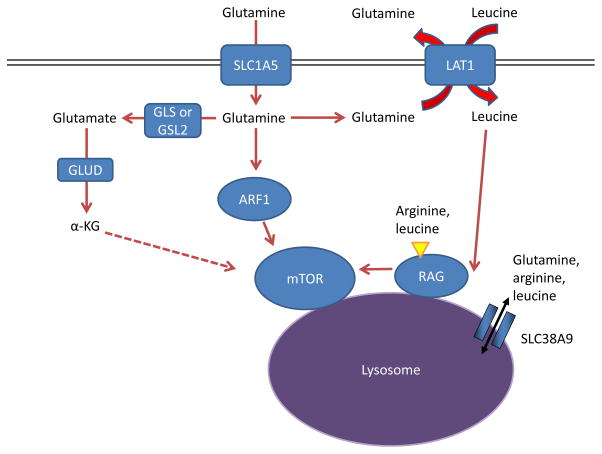
Amino acids stimulate the mTOR pathway, and amino acid pools rely on glutamine to
be maintained. Specifically, arginine and leucine are two amino acids that can
together almost fully stimulate mTOR complex 1 (mTORC1) through activation of
the RAS-related GTPase (RAG) complex, which in turn recruits mTORC1 to the
lysosome and stimulates its activity , , . Glutamine can contribute to
mTORC1 activation by being exchanged for essential amino acids, including
leucine, through the large neutral amino acid transporter 1 (LAT1; a heterodimer
of SLC7A5 and SLC3A2) transporter . This RAG-dependent regulation of mTOR is likely dependent
on the lysosomal amino acid transporter SLC38A9, which transports glutamine,
arginine, and leucine as substrates, , , as well as the leucine
sensor sestrin 2 (not shown in Figure) , . Although
the mechanism is not well understood, α-ketoglutarate (α-KG) may
regulate RAGB activity and mTOR activation downstream of glutamine metabolism
. Several
RAG-independent pathways of mTOR regulation by glutamine have also been
identified. Glutamine promotes mTOR localization to the lysosome (and thus
activity) through the RAS-family member ADP ribosylation factor 1 (ARF1) in a
poorly understood mechanism, as well as the TTT-RUVBL1/2 complex (not shown in
Figure) , . GLS, kidney-type glutaminase; GLS2,
liver-type glutaminase; GLUD, glutamate dehydrogenase.
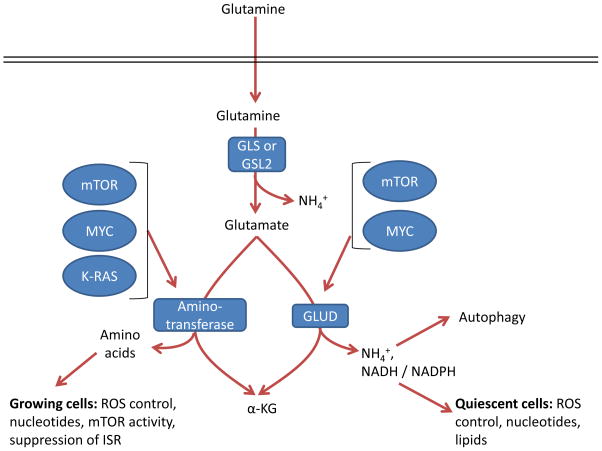
Glutamate can be converted by one of two different pathways into
α-ketoglutarate (α-KG), and the choice of which pathway is
influenced both by oncogene input and cell proliferation and metabolic state.
GLS, kidney-type glutaminase; GLS2, liver-type glutaminase; GLUD, glutamate
dehydrogenase; ISR, integrated stress response; ROS, reactive oxygen
species.
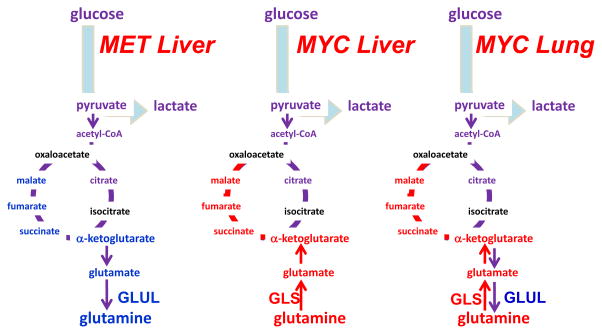
The oncoproteins MET and MYC lead to differing dependence on glutamine in
different cancer types, which is partially influenced by differential expression
of glutamine synthetase (GLUL) or glutaminase (GLS). α-KG,
α-ketoglutarate; OAA, oxaloacetate; Illustration is drawn from primary
data originally presented in Yuneva et al..
References
-
- Warburg O. On the origin of cancer cells. Science. 1956;123:309–14. - PubMed
-
- Lacey JM, Wilmore DW. Is glutamine a conditionally essential amino acid? Nutr Rev. 1990;48:297–309. - PubMed
-
- Rubin AL. Suppression of transformation by and growth adaptation to low concentrations of glutamine in NIH-3T3 cells. Cancer Res. 1990;50:2832–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
