

Backmasking in the yeast genome: encoding overlapping information for protein-coding and RNA degradation
- PMID: 27492286
- PMCID: PMC5041482
- DOI: 10.1093/nar/gkw683
Backmasking in the yeast genome: encoding overlapping information for protein-coding and RNA degradation
Abstract
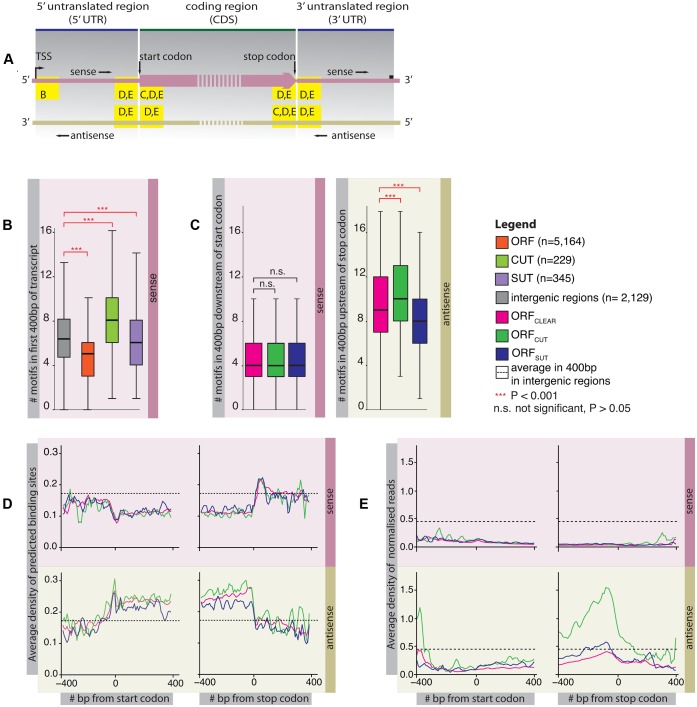
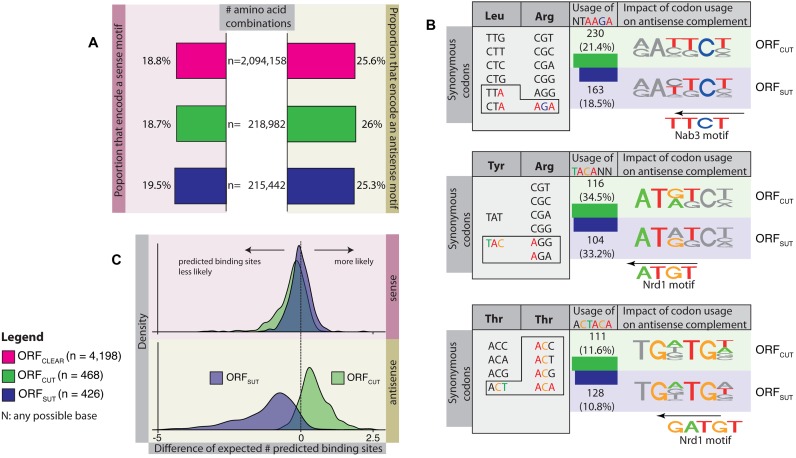
Backmasking is a recording technique used to hide a sound or message in a music track in reverse, meaning that it is only audible when the record is played backwards. Analogously, the compact yeast genome encodes for diverse sources of information such as overlapping coding and non-coding transcripts, and protein-binding sites on the two complementary DNA strands. Examples are the consensus binding site sequences of the RNA-binding proteins Nrd1 and Nab3 that target non-coding transcripts for degradation. Here, by examining the overlap of stable (SUTs, stable unannotated transcripts) and unstable (CUTs, cryptic unstable transcripts) transcripts with protein-coding genes, we show that the predicted Nrd1 and Nab3-binding site sequences occur at differing frequencies. They are always depleted in the sense direction of protein-coding genes, thus avoiding degradation of the transcript. However in the antisense direction, predicted binding sites occur at high frequencies in genes with overlapping unstable ncRNAs (CUTs), so limiting the availability of non-functional transcripts. In contrast they are depleted in genes with overlapping stable ncRNAs (SUTs), presumably to avoid degrading the non-coding transcript. The protein-coding genes maintain similar amino-acid contents, but they display distinct codon usages so that Nrd1 and Nab3-binding sites can arise at differing frequencies in antisense depending on the overlapping transcript type. Our study demonstrates how yeast has evolved to encode multiple layers of information-protein-coding genes in one strand and the relative chance of degrading antisense RNA in the other strand-in the same regions of a compact genome.
© The Author(s) 2016. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Camblong J., Iglesias N., Fickentscher C., Dieppois G., Stutz F. Antisense RNA stabilization induces transcriptional gene silencing via histone deacetylation in S. cerevisiae. Cell. 2007;131:706–717. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
