

PTEN regulates EG5 to control spindle architecture and chromosome congression during mitosis
- PMID: 27492783
- PMCID: PMC4980451
- DOI: 10.1038/ncomms12355
PTEN regulates EG5 to control spindle architecture and chromosome congression during mitosis
Abstract
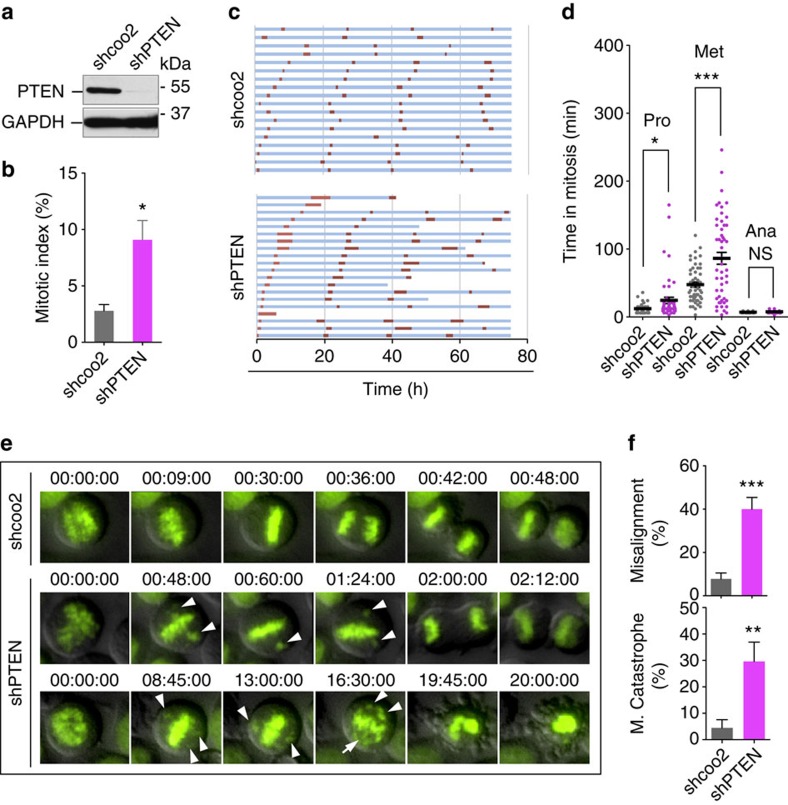
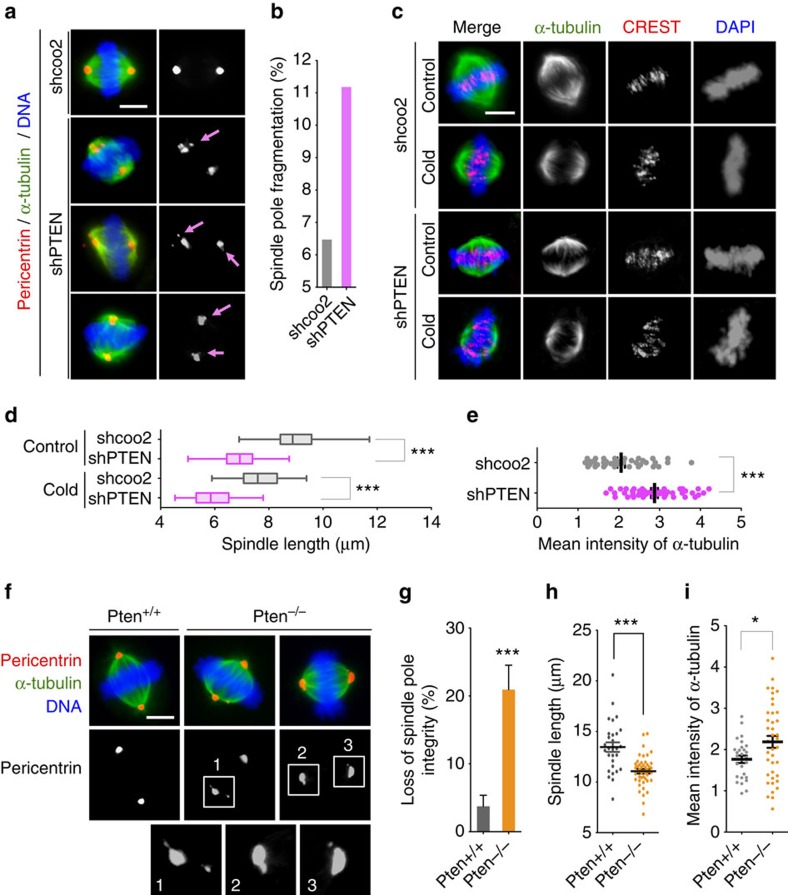
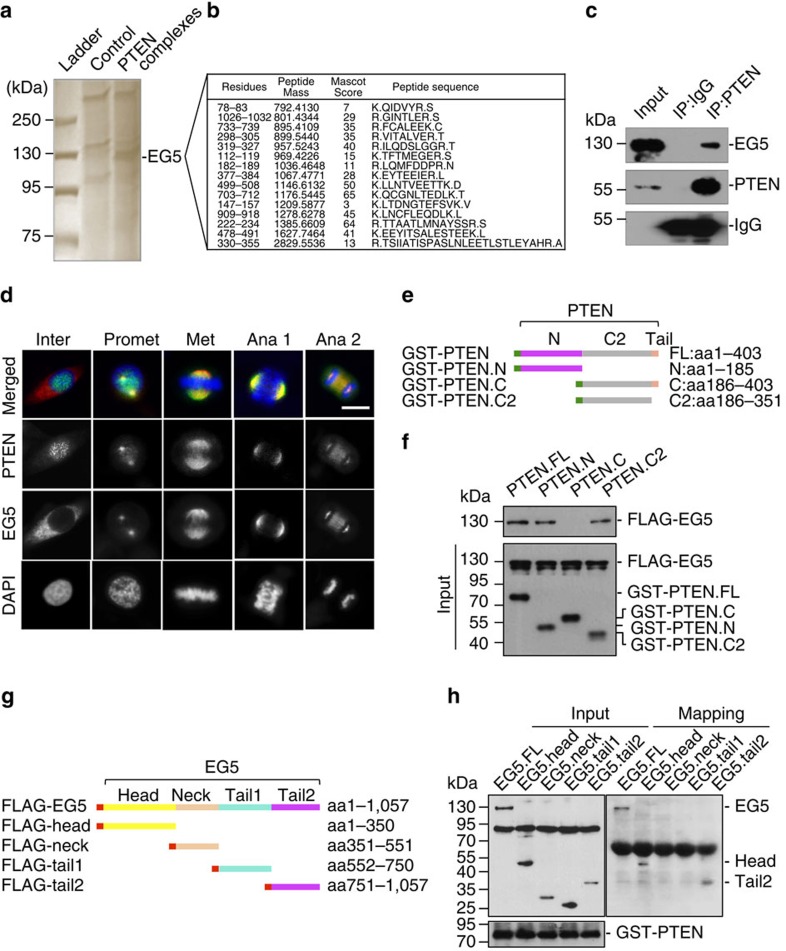
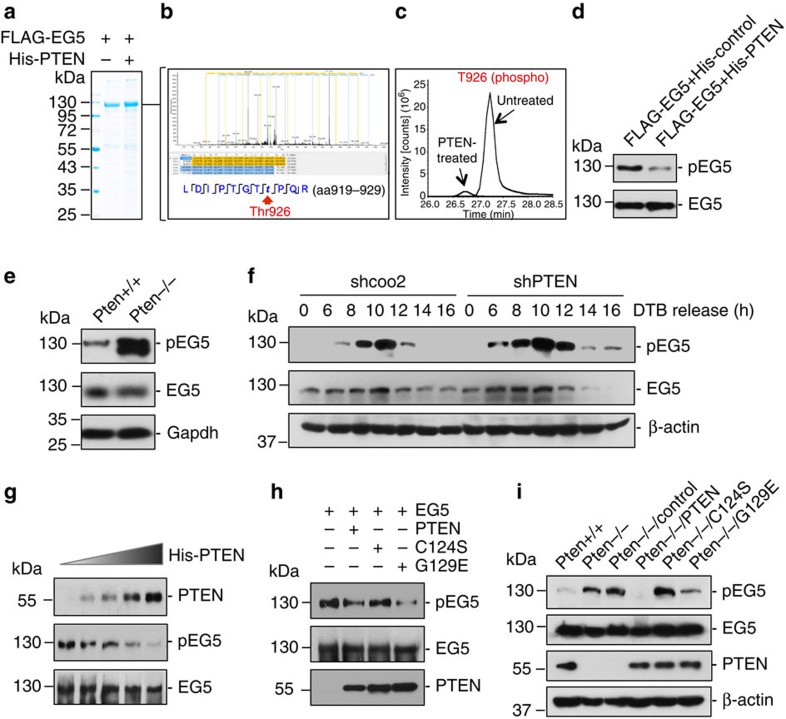
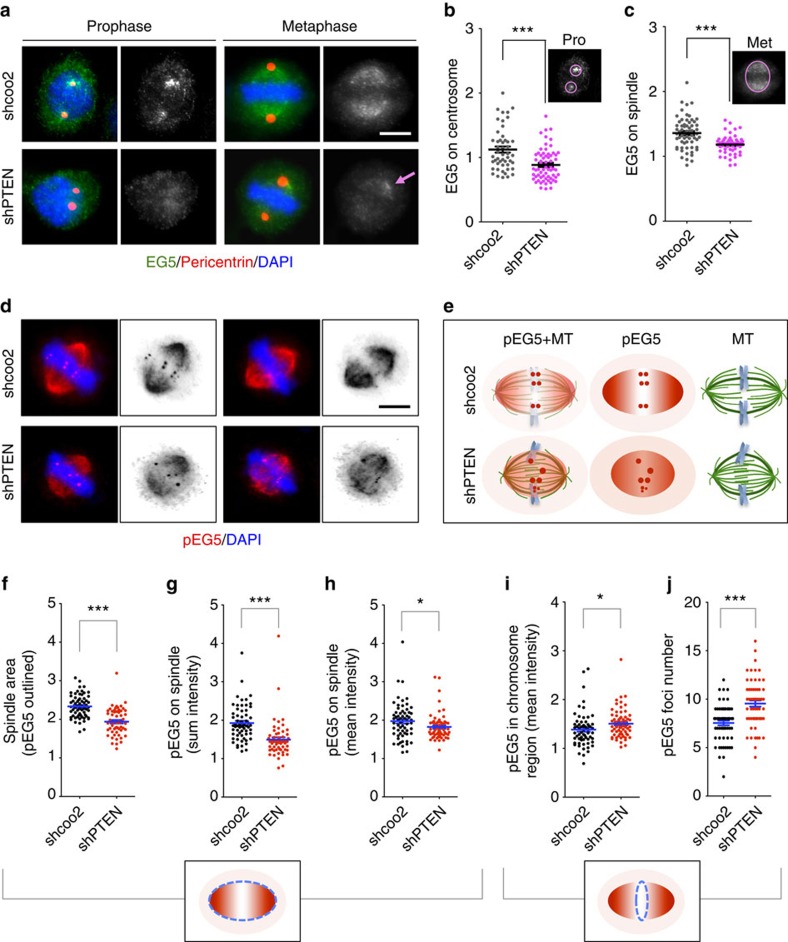
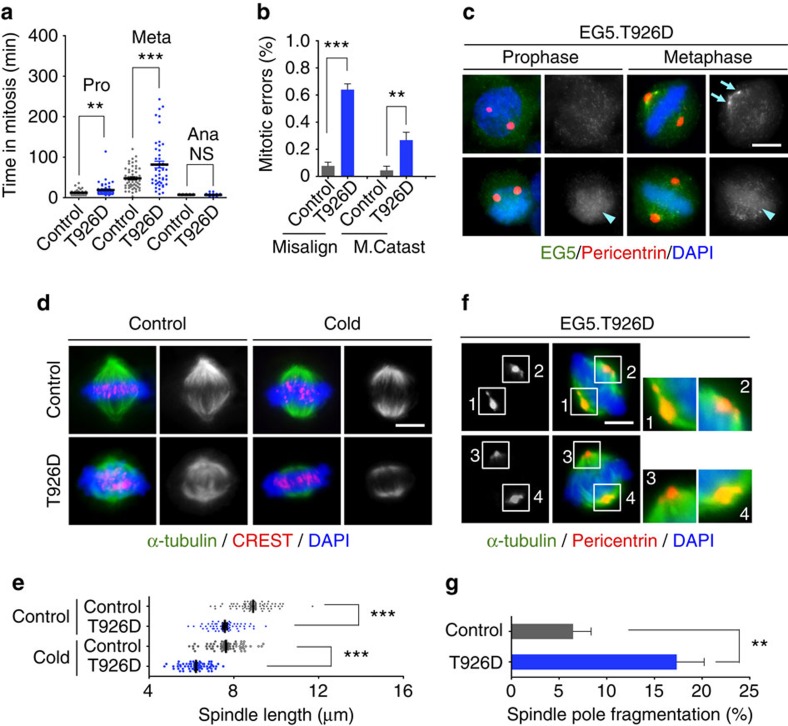
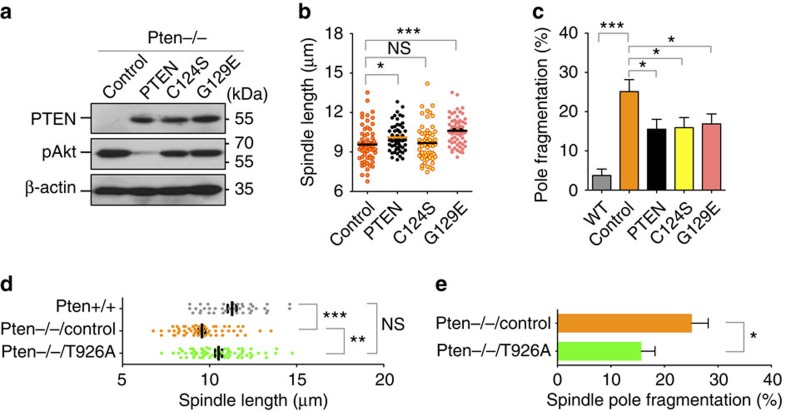
Architectural integrity of the mitotic spindle is required for efficient chromosome congression and accurate chromosome segregation to ensure mitotic fidelity. Tumour suppressor PTEN has multiple functions in maintaining genome stability. Here we report an essential role of PTEN in mitosis through regulation of the mitotic kinesin motor EG5 for proper spindle architecture and chromosome congression. PTEN depletion results in chromosome misalignment in metaphase, often leading to catastrophic mitotic failure. In addition, metaphase cells lacking PTEN exhibit defects of spindle geometry, manifested prominently by shorter spindles. PTEN is associated and co-localized with EG5 during mitosis. PTEN deficiency induces aberrant EG5 phosphorylation and abrogates EG5 recruitment to the mitotic spindle apparatus, leading to spindle disorganization. These data demonstrate the functional interplay between PTEN and EG5 in controlling mitotic spindle structure and chromosome behaviour during mitosis. We propose that PTEN functions to equilibrate mitotic phosphorylation for proper spindle formation and faithful genomic transmission.
Figures
Similar articles
-
PTEN in the maintenance of genome integrity: From DNA replication to chromosome segregation.Bioessays. 2017 Oct;39(10):10.1002/bies.201700082. doi: 10.1002/bies.201700082. Epub 2017 Sep 11. Bioessays. 2017. PMID: 28891157 Free PMC article. Review.
-
UHRF1 promotes spindle assembly and chromosome congression by catalyzing EG5 polyubiquitination.J Cell Biol. 2023 Nov 6;222(11):e202210093. doi: 10.1083/jcb.202210093. Epub 2023 Sep 20. J Cell Biol. 2023. PMID: 37728657 Free PMC article.
-
Protein Phosphatase 2A (PP2A) Regulates EG5 to Control Mitotic Progression.Sci Rep. 2017 May 9;7(1):1630. doi: 10.1038/s41598-017-01915-w. Sci Rep. 2017. PMID: 28487562 Free PMC article.
-
Pten regulates spindle pole movement through Dlg1-mediated recruitment of Eg5 to centrosomes.Nat Cell Biol. 2016 Jul;18(7):814-21. doi: 10.1038/ncb3369. Epub 2016 May 30. Nat Cell Biol. 2016. PMID: 27240320 Free PMC article.
-
Non-canonical functions of the mitotic kinesin Eg5.Thorac Cancer. 2018 Aug;9(8):904-910. doi: 10.1111/1759-7714.12792. Epub 2018 Jun 21. Thorac Cancer. 2018. PMID: 29927078 Free PMC article. Review.
Cited by
-
Phosphorylation of XPD drives its mitotic role independently of its DNA repair and transcription functions.Sci Adv. 2022 Aug 19;8(33):eabp9457. doi: 10.1126/sciadv.abp9457. Epub 2022 Aug 17. Sci Adv. 2022. PMID: 35977011 Free PMC article.
-
Potential Roles of PTEN on Longevity in Two Closely Related Argopecten Scallops With Distinct Lifespans.Front Physiol. 2022 Jul 12;13:872562. doi: 10.3389/fphys.2022.872562. eCollection 2022. Front Physiol. 2022. PMID: 35903068 Free PMC article.
-
PTEN in the maintenance of genome integrity: From DNA replication to chromosome segregation.Bioessays. 2017 Oct;39(10):10.1002/bies.201700082. doi: 10.1002/bies.201700082. Epub 2017 Sep 11. Bioessays. 2017. PMID: 28891157 Free PMC article. Review.
-
PTEN Protein Phosphatase Activity Is Not Required for Tumour Suppression in the Mouse Prostate.Biomolecules. 2022 Oct 19;12(10):1511. doi: 10.3390/biom12101511. Biomolecules. 2022. PMID: 36291720 Free PMC article.
-
Unmasking PTEN's nuclear functions.Cell Cycle. 2016 Dec 16;15(24):3341-3342. doi: 10.1080/15384101.2016.1241606. Epub 2016 Oct 20. Cell Cycle. 2016. PMID: 27764547 Free PMC article. No abstract available.
References
-
- Yin Y. & Shen W. H. PTEN: a new guardian of the genome. Oncogene 27, 5443–5453 (2008). - PubMed
-
- Puc J. et al.. Lack of PTEN sequesters CHK1 and initiates genetic instability. Cancer Cell 7, 193–204 (2005). - PubMed
-
- Shen W. H. et al.. Essential role for nuclear PTEN in maintaining chromosomal integrity. Cell 128, 157–170 (2007). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
