

Kinetic and thermodynamic framework for P4-P6 RNA reveals tertiary motif modularity and modulation of the folding preferred pathway
- PMID: 27493222
- PMCID: PMC5003260
- DOI: 10.1073/pnas.1525082113
Kinetic and thermodynamic framework for P4-P6 RNA reveals tertiary motif modularity and modulation of the folding preferred pathway
Abstract
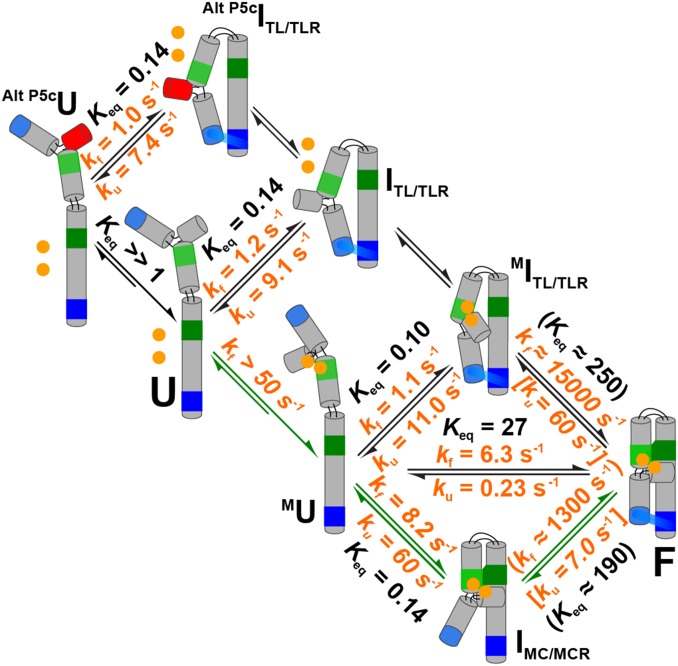
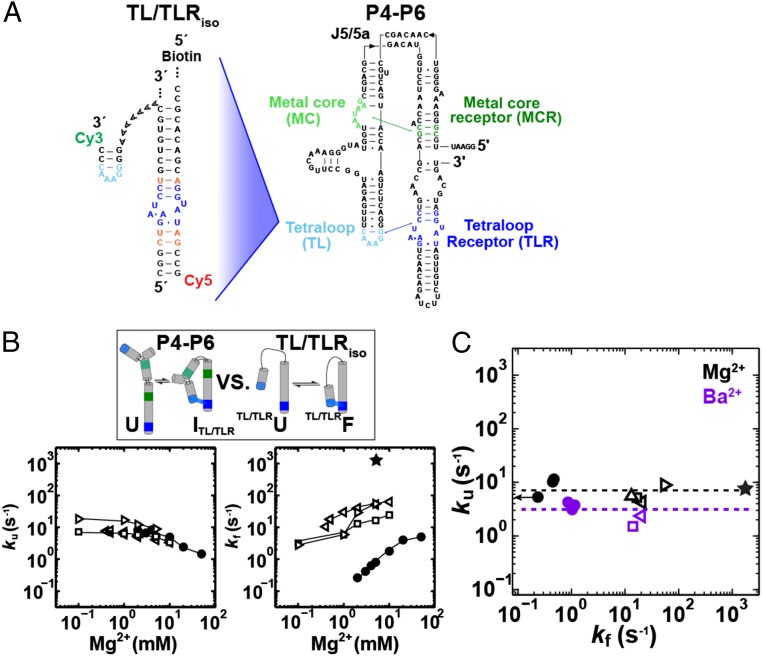
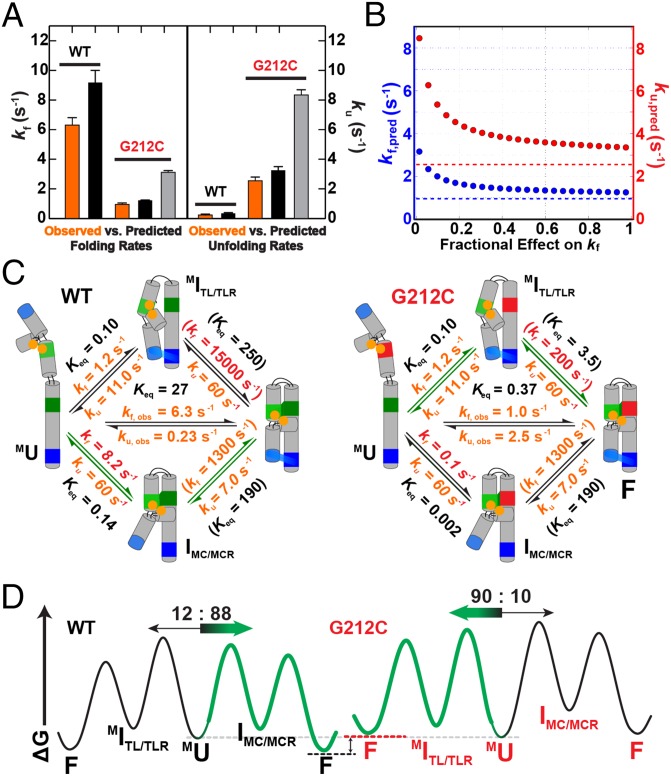
The past decade has seen a wealth of 3D structural information about complex structured RNAs and identification of functional intermediates. Nevertheless, developing a complete and predictive understanding of the folding and function of these RNAs in biology will require connection of individual rate and equilibrium constants to structural changes that occur in individual folding steps and further relating these steps to the properties and behavior of isolated, simplified systems. To accomplish these goals we used the considerable structural knowledge of the folded, unfolded, and intermediate states of P4-P6 RNA. We enumerated structural states and possible folding transitions and determined rate and equilibrium constants for the transitions between these states using single-molecule FRET with a series of mutant P4-P6 variants. Comparisons with simplified constructs containing an isolated tertiary contact suggest that a given tertiary interaction has a stereotyped rate for breaking that may help identify structural transitions within complex RNAs and simplify the prediction of folding kinetics and thermodynamics for structured RNAs from their parts. The preferred folding pathway involves initial formation of the proximal tertiary contact. However, this preference was only ∼10 fold and could be reversed by a single point mutation, indicating that a model akin to a protein-folding contact order model will not suffice to describe RNA folding. Instead, our results suggest a strong analogy with a modified RNA diffusion-collision model in which tertiary elements within preformed secondary structures collide, with the success of these collisions dependent on whether the tertiary elements are in their rare binding-competent conformations.
Keywords: RNA folding; RNA tertiary motifs; folding pathways; kinetics; single-molecule FRET.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
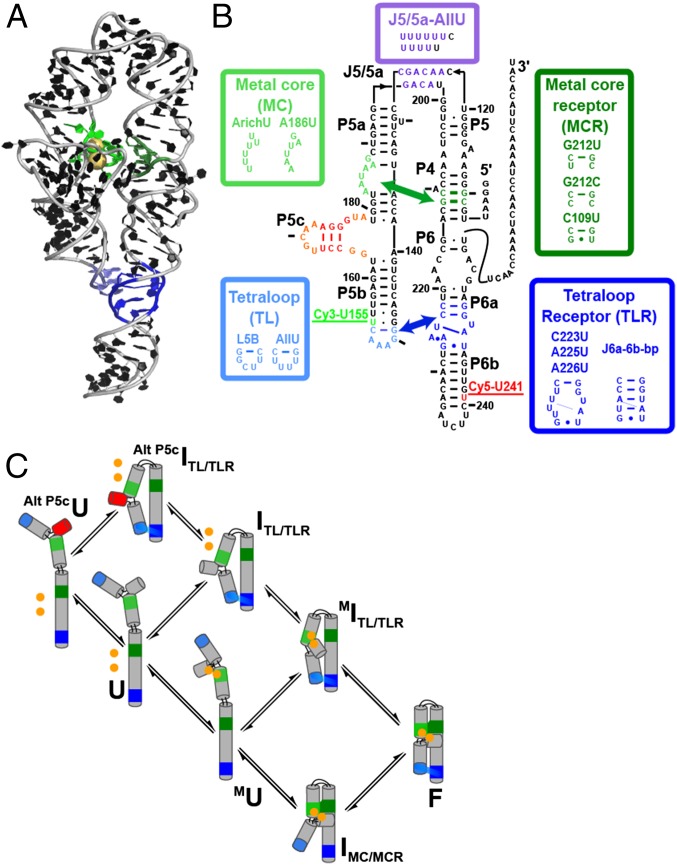
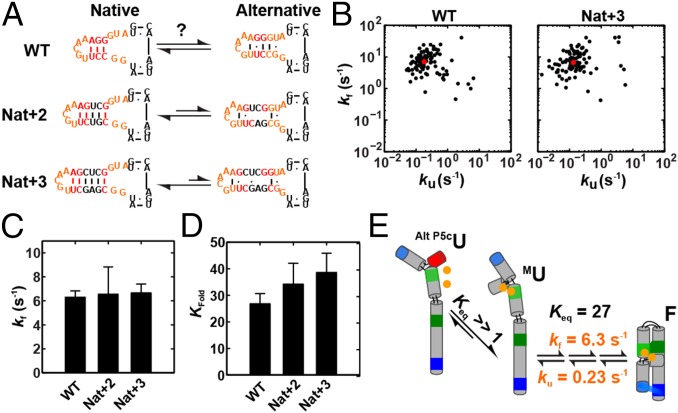
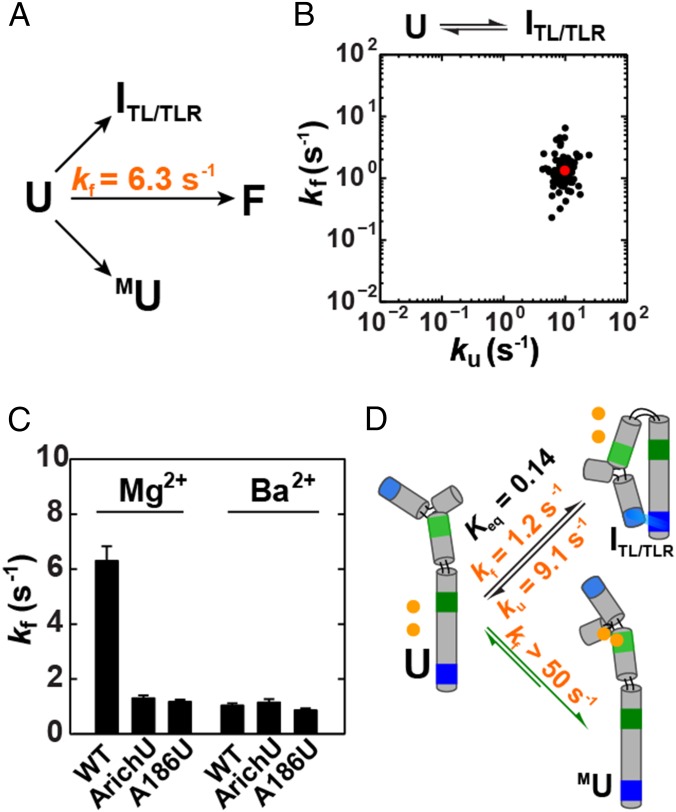
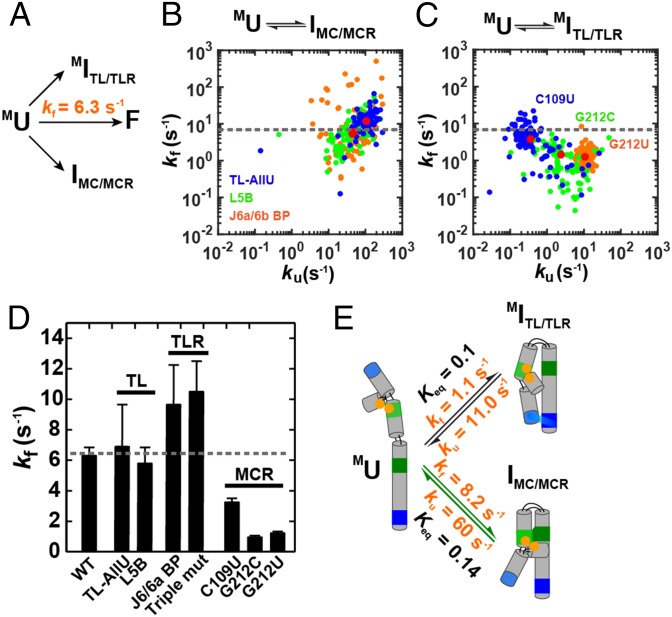
References
-
- Mercer TR, Mattick JS. Structure and function of long noncoding RNAs in epigenetic regulation. Nat Struct Mol Biol. 2013;20(3):300–307. - PubMed
-
- Cech TR, Steitz JA. The noncoding RNA revolution-trashing old rules to forge new ones. Cell. 2014;157(1):77–94. - PubMed
-
- Staley JP, Guthrie C. Mechanical devices of the spliceosome: Motors, clocks, springs, and things. Cell. 1998;92(3):315–326. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
