

The Tumor Microenvironment Represses T Cell Mitochondrial Biogenesis to Drive Intratumoral T Cell Metabolic Insufficiency and Dysfunction
- PMID: 27496732
- PMCID: PMC5207350
- DOI: 10.1016/j.immuni.2016.07.009
The Tumor Microenvironment Represses T Cell Mitochondrial Biogenesis to Drive Intratumoral T Cell Metabolic Insufficiency and Dysfunction
Erratum in
-
The Tumor Microenvironment Represses T Cell Mitochondrial Biogenesis to Drive Intratumoral T Cell Metabolic Insufficiency and Dysfunction.Immunity. 2016 Sep 20;45(3):701-703. doi: 10.1016/j.immuni.2016.08.009. Epub 2016 Sep 20. Immunity. 2016. PMID: 27653602 No abstract available.
Abstract
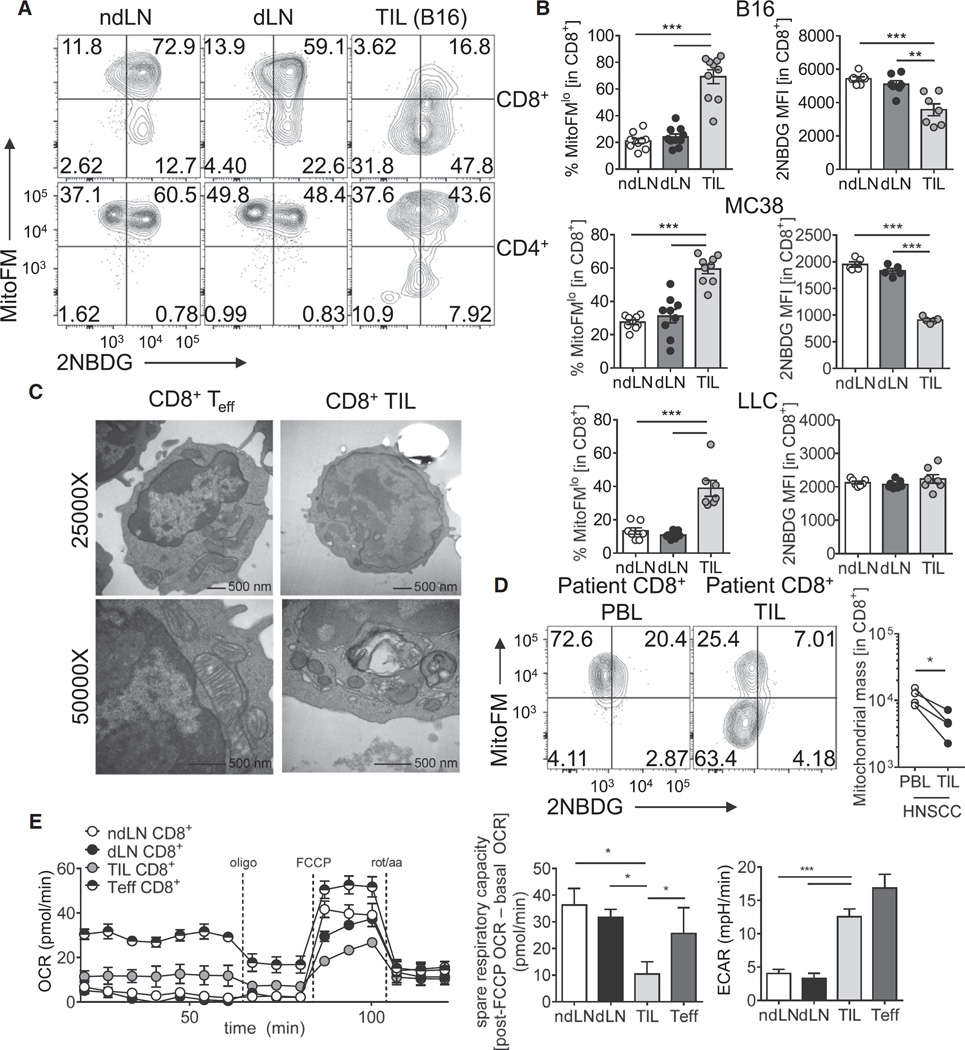
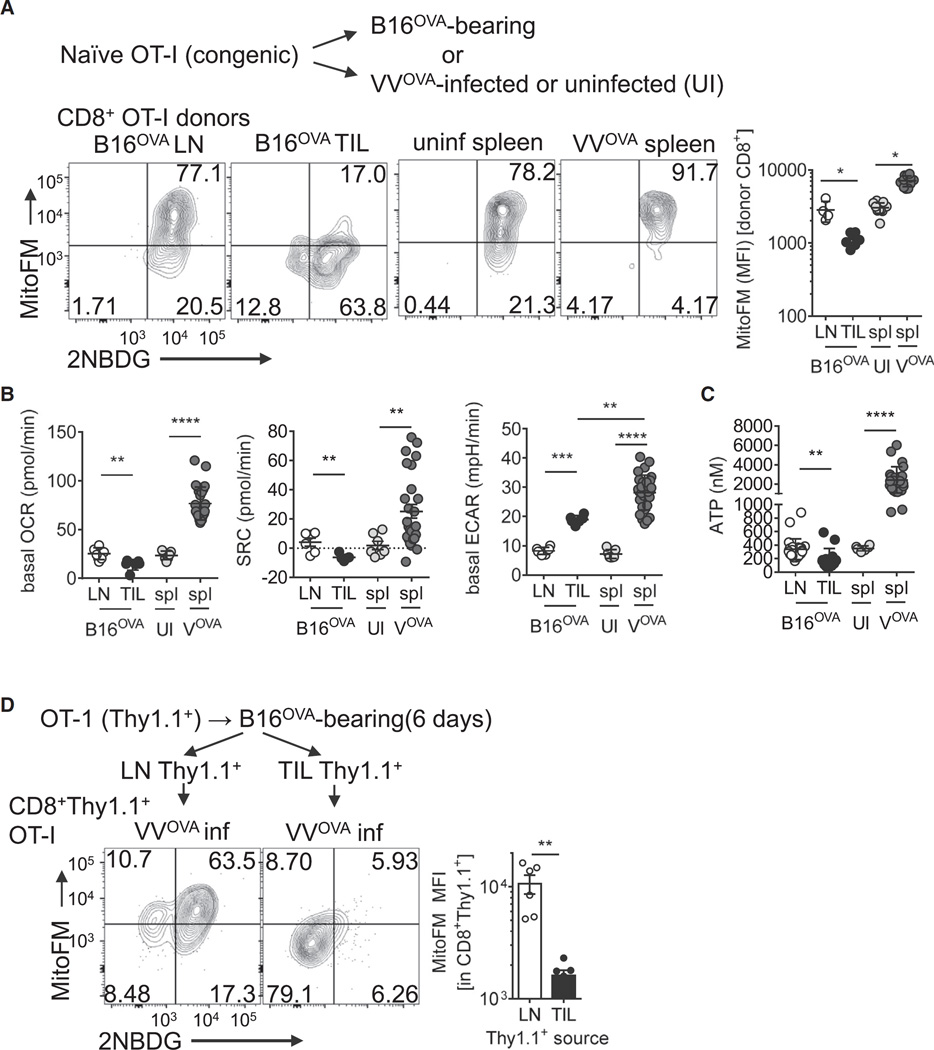
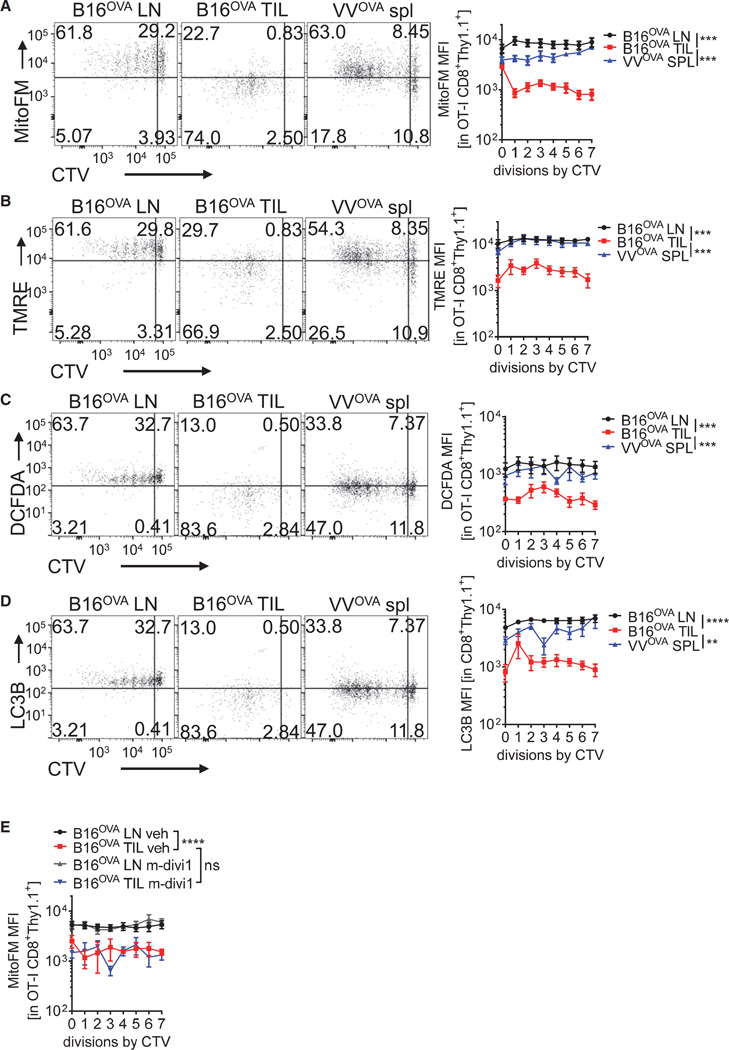
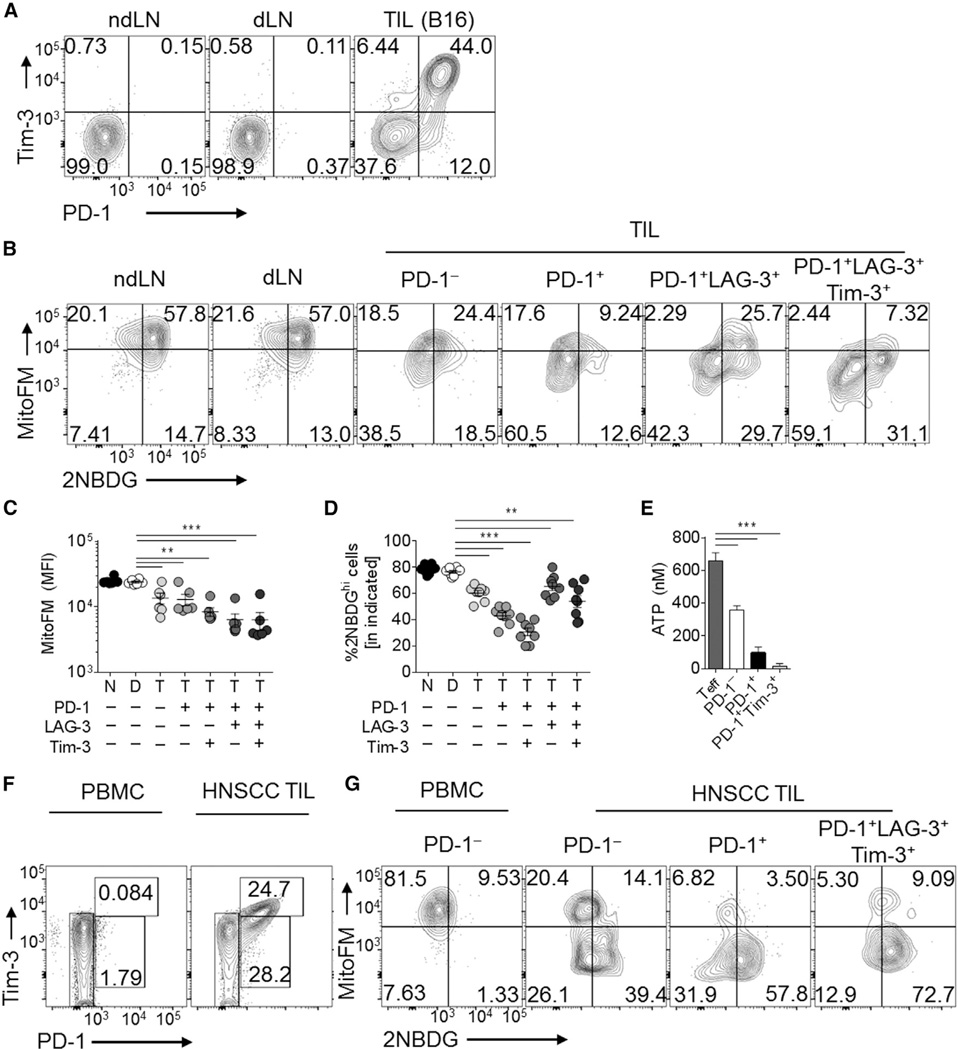
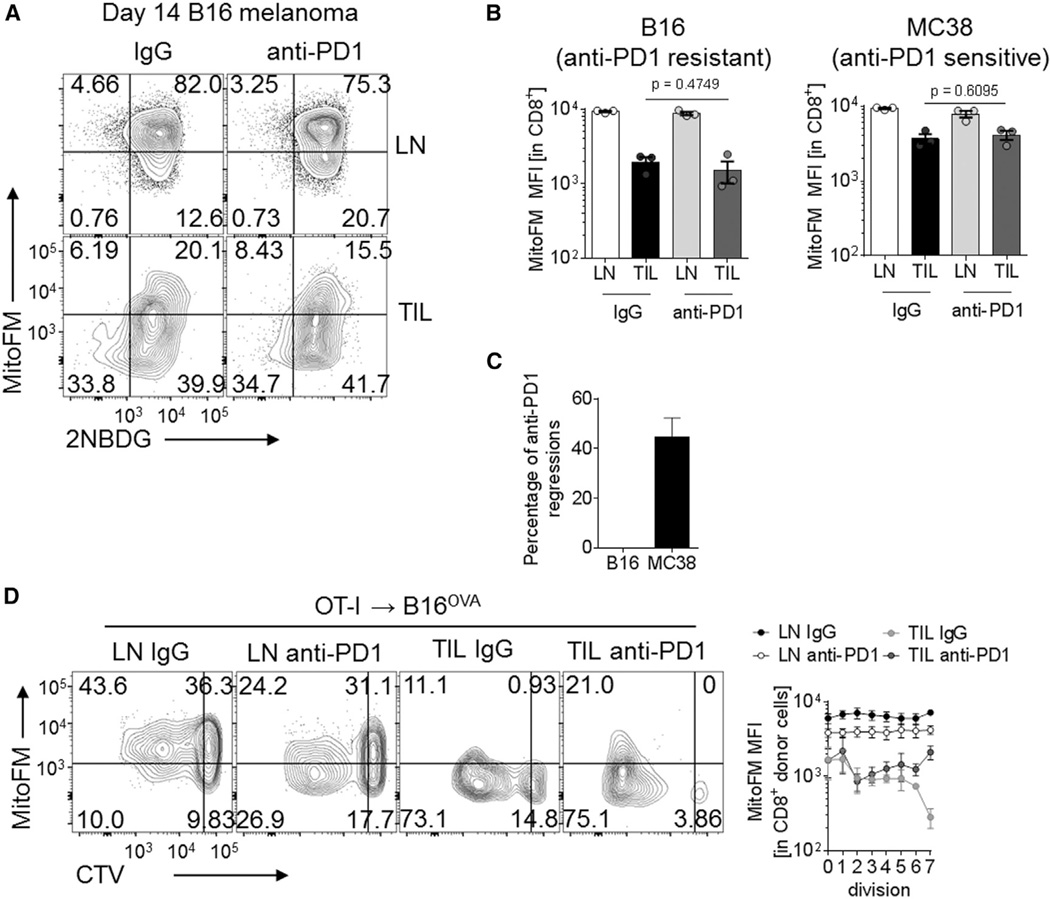
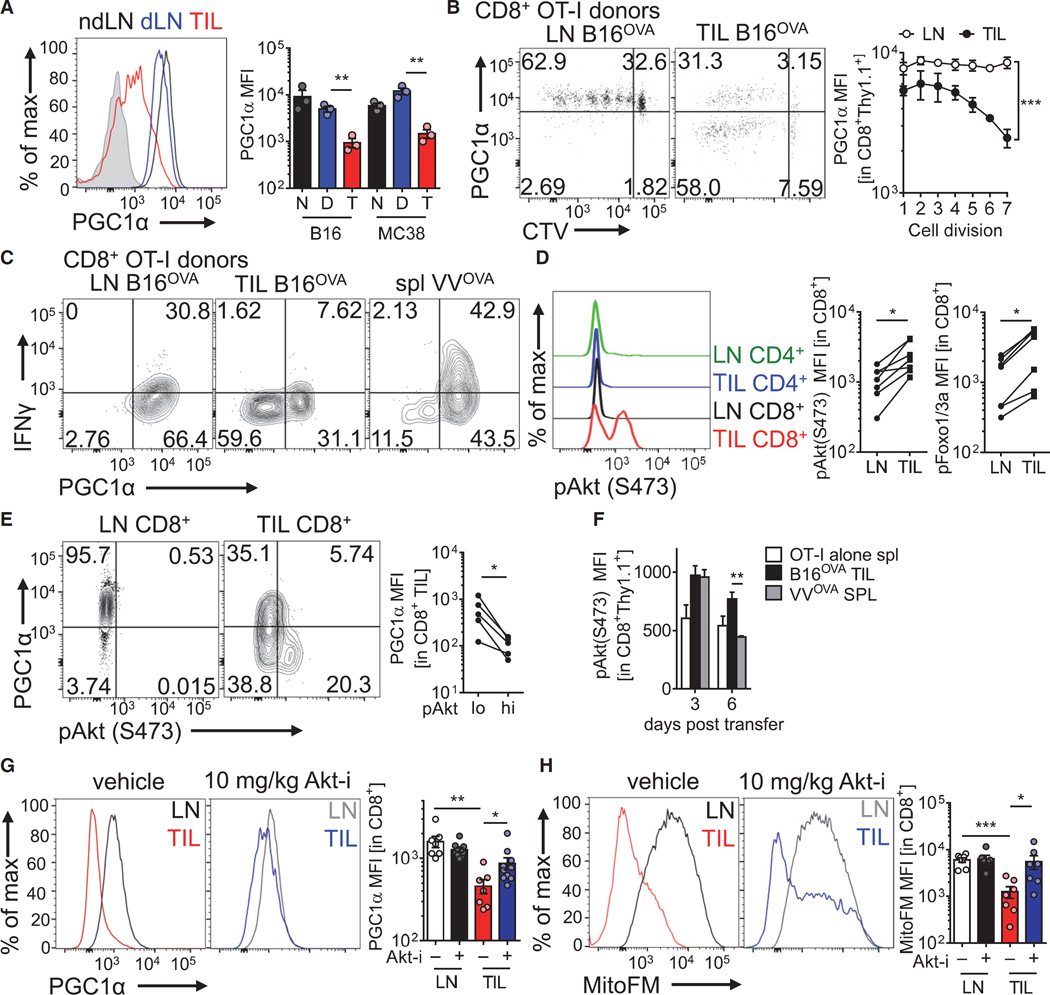
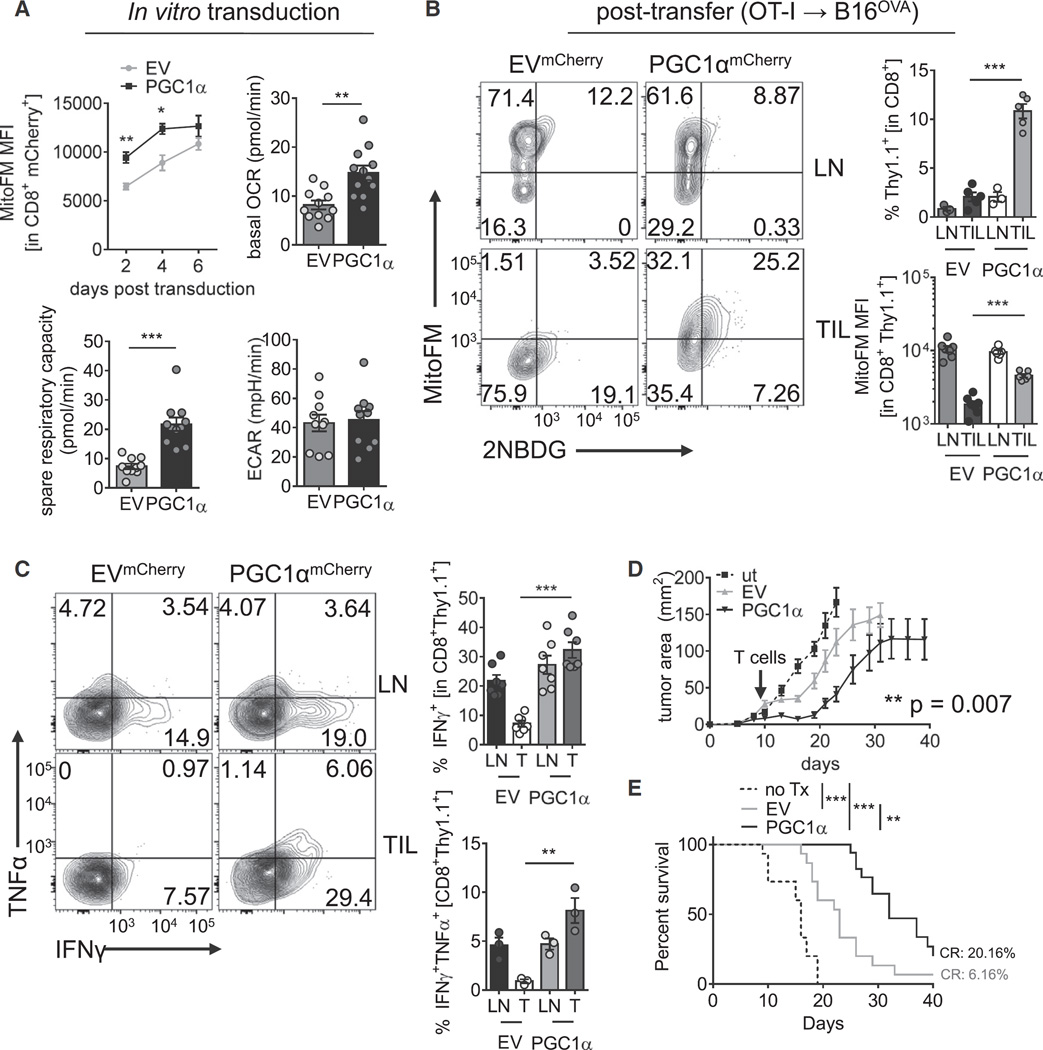
Although tumor-specific T cells recognize cancer cells, they are often rendered dysfunctional due to an immunosuppressive microenvironment. Here we showed that T cells demonstrated persistent loss of mitochondrial function and mass when infiltrating murine and human tumors, an effect specific to the tumor microenvironment and not merely caused by activation. Tumor-infiltrating T cells showed a progressive loss of PPAR-gamma coactivator 1α (PGC1α), which programs mitochondrial biogenesis, induced by chronic Akt signaling in tumor-specific T cells. Reprogramming tumor-specific T cells through enforced expression of PGC1α resulted in superior intratumoral metabolic and effector function. Our data support a model in which signals in the tumor microenvironment repress T cell oxidative metabolism, resulting in effector cells with metabolic needs that cannot be met. Our studies also suggest that modulation or reprogramming of the altered metabolism of tumor-infiltrating T cells might represent a potential strategy to reinvigorate dysfunctional T cells for cancer treatment.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Feeling Worn Out? PGC1α to the Rescue for Dysfunctional Mitochondria in T Cell Exhaustion.Immunity. 2016 Aug 16;45(2):233-5. doi: 10.1016/j.immuni.2016.07.024. Immunity. 2016. PMID: 27533009
-
T Cell responses: Defective mitochondria disrupt CD8(+) T cells.Nat Rev Immunol. 2016 Aug 25;16(9):534-5. doi: 10.1038/nri.2016.98. Nat Rev Immunol. 2016. PMID: 27558410 No abstract available.
References
-
- Alvarez-Guardia D, Palomer X, Coll T, Davidson MM, Chan TO, Feldman AM, Laguna JC, Vázquez-Carrera M. The p65 subunit of NF-kappaB binds to PGC-1alpha, linking inflammation and metabolic disturbances in cardiac cells. Cardiovasc. Res. 2010;87:449–458. - PubMed
-
- Cottet-Rousselle C, Ronot X, Leverve X, Mayol J-F. Cytometric assessment of mitochondria using fluorescent probes. Cytometry A. 2011;79:405–425. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
