

Mitochondrial DNA depletion causes decreased ROS production and resistance to apoptosis
- PMID: 27499009
- PMCID: PMC5029958
- DOI: 10.3892/ijmm.2016.2697
Mitochondrial DNA depletion causes decreased ROS production and resistance to apoptosis
Abstract
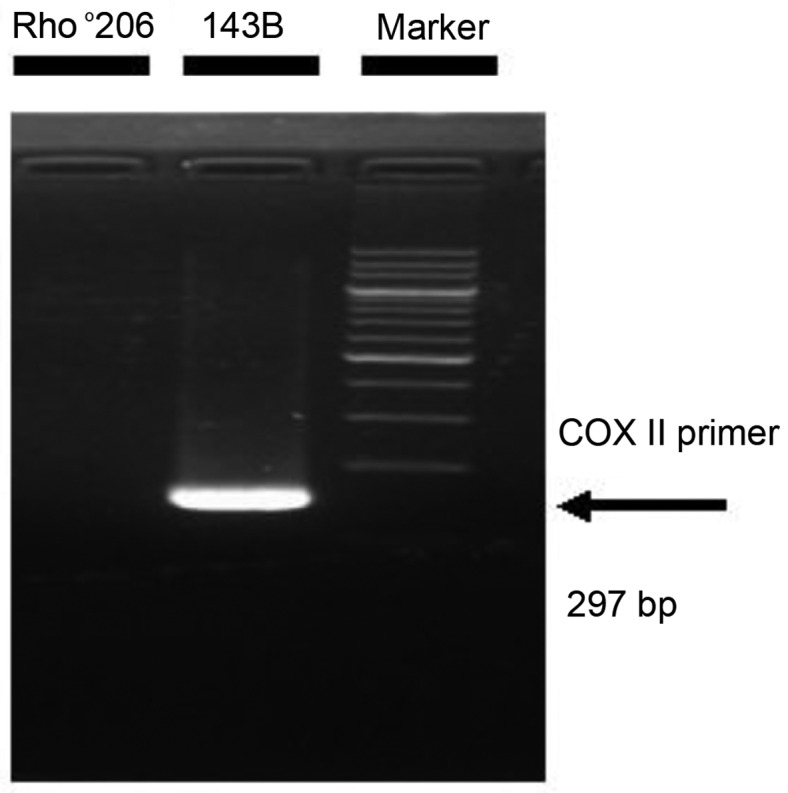
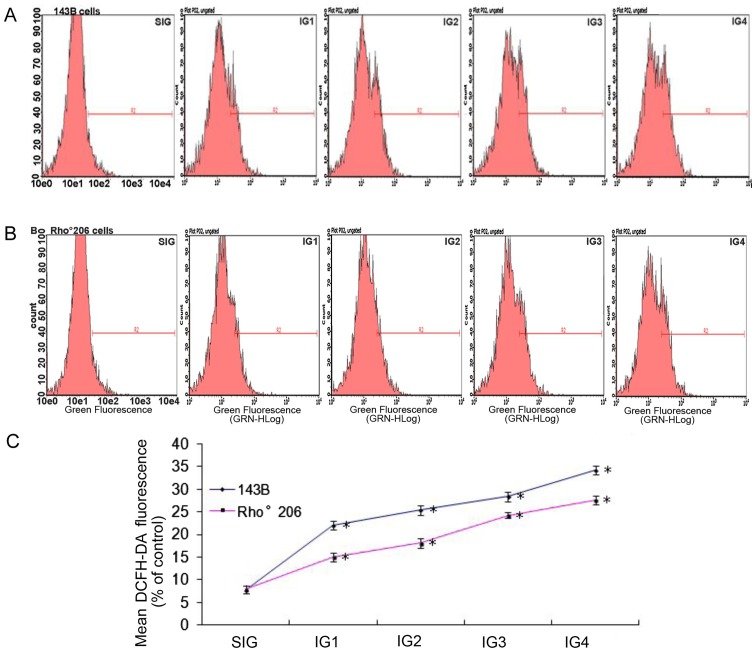
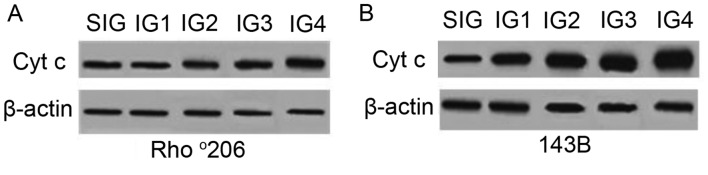
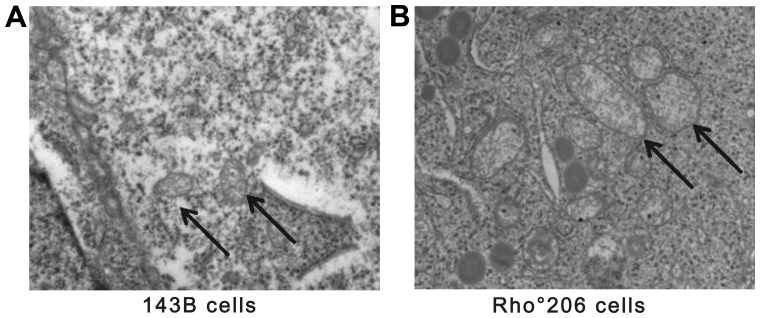
Mitochondrial DNA (mtDNA) depletion occurs frequently in many diseases including cancer. The present study was designed in order to examine the hypothesis that mtDNA‑depleted cells are resistant to apoptosis and to explore the possible mechanisms responsible for this effect. Parental human osteosarcoma 143B cells and mtDNA‑deficient (Rho˚ or ρ˚) 206 cells (derived from 143B cells) were exposed to different doses of solar-simulated ultraviolet (UV) radiation. The effects of solar irradiation on cell morphology were observed under both light and fluorescence microscopes. Furthermore, apoptosis, mitochondrial membrane potential (MMP) disruption and reactive oxygen species (ROS) production were detected and measured by flow cytometry. In both cell lines, apoptosis and ROS production were clearly increased, whereas MMP was slightly decreased. However, apoptosis and ROS production were reduced in the Rho˚206 cells compared with the 143B cells. We also performed western blot analysis and demonstrated the increased release of cytosolic Cyt c from mitochondria in the 143B cells compared with that in the Rho˚206 cells. Thus, we concluded that Rho˚206 cells exhibit more resistance to solar‑simulated UV radiation‑induced apoptosis at certain doses than 143B cells and this is possibly due to decreased ROS production.
Figures


References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
