

Targeted alternative splicing of TAF4: a new strategy for cell reprogramming
- PMID: 27499390
- PMCID: PMC4976350
- DOI: 10.1038/srep30852
Targeted alternative splicing of TAF4: a new strategy for cell reprogramming
Abstract
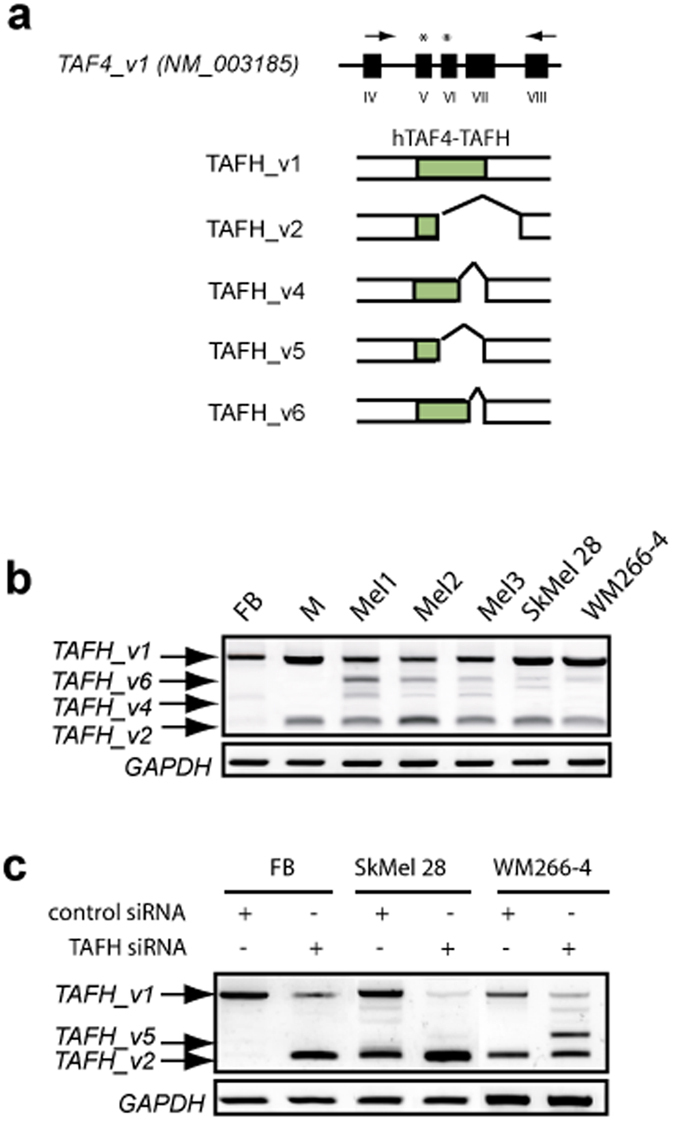
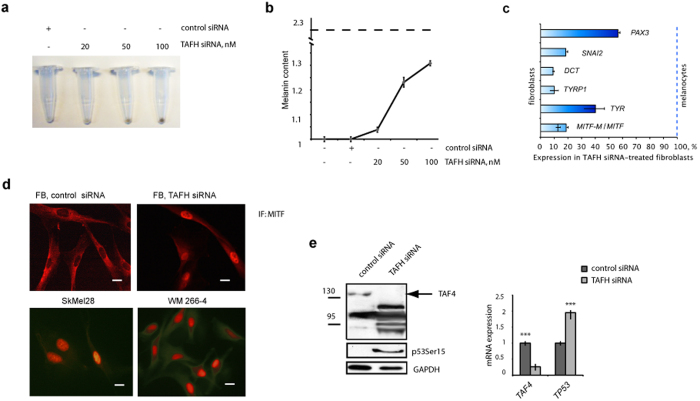
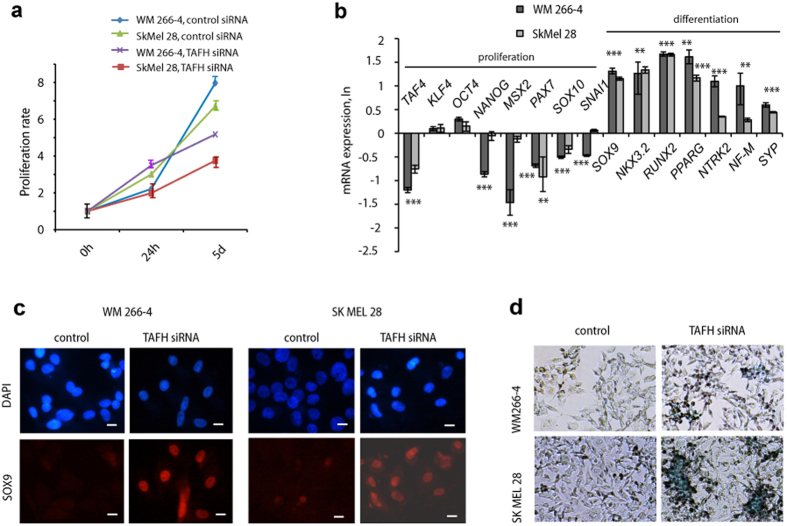
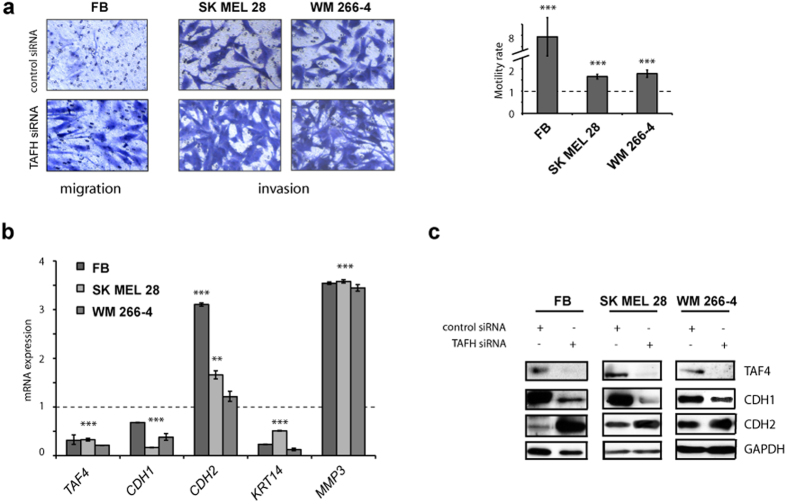
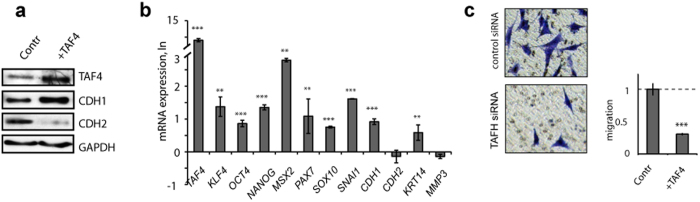
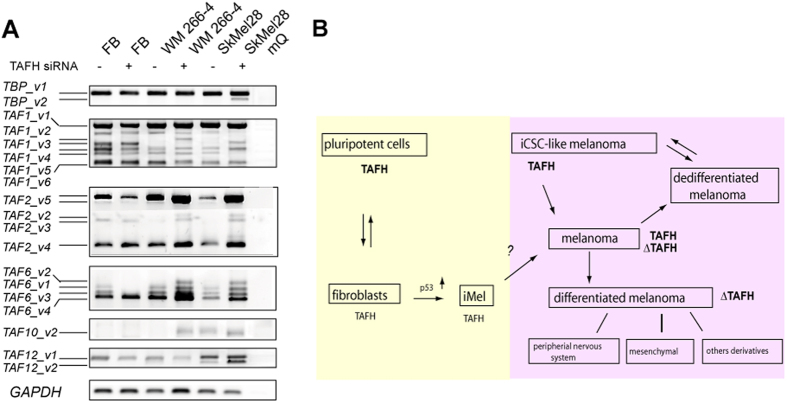
Reprogramming of somatic cells has become a versatile tool for biomedical research and for regenerative medicine. In the current study, we show that manipulating alternative splicing (AS) is a highly potent strategy to produce cells for therapeutic applications. We demonstrate that silencing of hTAF4-TAFH activity of TAF4 converts human facial dermal fibroblasts to melanocyte-like (iMel) cells. iMel cells produce melanin and express microphthalmia-associated transcription factor (MITF) and its target genes at levels comparable to normal melanocytes. Reprogramming of melanoma cells by manipulation with hTAF4-TAFH activity upon TAFH RNAi enforces cell differentiation towards chondrogenic pathway, whereas ectoptic expression of TAF4 results in enhanced multipotency and neural crest-like features in melanoma cells. In both cell states, iMels and cancer cells, hTAF4-TAFH activity controls migration by supporting E- to N-cadherin switches. From our data, we conclude that targeted splicing of hTAF4-TAFH coordinates AS of other TFIID subunits, underscoring the role of TAF4 in synchronised changes of Pol II complex composition essential for efficient cellular reprogramming. Taken together, targeted AS of TAF4 provides a unique strategy for generation of iMels and recapitulating stages of melanoma progression.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
